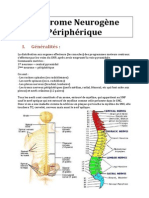
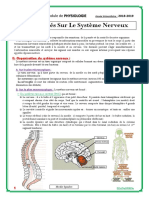
Vous aimerez peut-être aussi
- Essai de topographie cérébrale par la cérébrotomie méthodique: Conservation des pièces normales et pathologiques par un procédé particulierD'EverandEssai de topographie cérébrale par la cérébrotomie méthodique: Conservation des pièces normales et pathologiques par un procédé particulierPas encore d'évaluation
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- 3 - Moelle, Organe ConductionDocument15 pages3 - Moelle, Organe ConductionEmelie MelodyPas encore d'évaluation
- La Moelle EpiniereDocument13 pagesLa Moelle EpiniereMOUSTAPHAPas encore d'évaluation
- SOMESTHESIE-Document12 pagesSOMESTHESIE-mbouillesissoko55Pas encore d'évaluation
- 8 - Somesthesie Dr. LassakeurDocument38 pages8 - Somesthesie Dr. LassakeurAmmar ChaouiPas encore d'évaluation
- Physio SOMESTHESIEDocument25 pagesPhysio SOMESTHESIEEsperancePas encore d'évaluation
- Les Fonctions SomésthésiquesDocument7 pagesLes Fonctions Somésthésiquesmohamed1539Pas encore d'évaluation
- SEQ 4-COURS Redige-Comportement-ReflexeDocument6 pagesSEQ 4-COURS Redige-Comportement-ReflexeLadetPas encore d'évaluation
- Les Racines RachidiennesDocument3 pagesLes Racines RachidiennesSaken667Pas encore d'évaluation
- 2-La Moelle SpinaleDocument9 pages2-La Moelle SpinalebPas encore d'évaluation
- 09 Neurologie - Sémio Sandoz PDFDocument50 pages09 Neurologie - Sémio Sandoz PDFLaura PussPas encore d'évaluation
- Fichier Produit 2337 PDFDocument13 pagesFichier Produit 2337 PDFHadjer SebihiPas encore d'évaluation
- SomesthesieDocument10 pagesSomesthesieasma2490Pas encore d'évaluation
- Systématisation de La Moelle ÉpinièreDocument12 pagesSystématisation de La Moelle ÉpinièreTala ToroPas encore d'évaluation
- Neuro D1Document26 pagesNeuro D1Clément BologniniPas encore d'évaluation
- UE3 - Physiologie - Neurophysiologie 2Document14 pagesUE3 - Physiologie - Neurophysiologie 2sun-nee-chan9Pas encore d'évaluation
- La Physiologie Du Systeme Nerveux Centrale DR GuedjatiDocument33 pagesLa Physiologie Du Systeme Nerveux Centrale DR GuedjatiMedMan6100% (2)
- CM NEUROSCIENCES (1)Document11 pagesCM NEUROSCIENCES (1)MassePas encore d'évaluation
- Moelle EpiniereDocument6 pagesMoelle Epiniereariane.vasseur29Pas encore d'évaluation
- 3 SomesthésieDocument25 pages3 SomesthésieSahouin Claude MedardPas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Chapitre 2 MOUV201Document20 pagesChapitre 2 MOUV201Matéo ROBINPas encore d'évaluation
- 5 - Physiologie de La SomésthésieDocument11 pages5 - Physiologie de La SomésthésieEmelie MelodyPas encore d'évaluation
- Physiopath3an16-10douleur ArkounDocument29 pagesPhysiopath3an16-10douleur ArkounEbePas encore d'évaluation
- La SomesthésieDocument3 pagesLa SomesthésieZied Bouabene100% (1)
- Anatomie ComplèteDocument118 pagesAnatomie ComplèteZakaria ChohoPas encore d'évaluation
- Tissus MusculairesDocument16 pagesTissus MusculairesandriePas encore d'évaluation
- ANATOMIE SN 2024Document53 pagesANATOMIE SN 2024lwakwanasaidesPas encore d'évaluation
- Weber - Système Nerveux VégétatifDocument5 pagesWeber - Système Nerveux VégétatifbureljulienPas encore d'évaluation
- SyndromeDocument19 pagesSyndromeenimaPas encore d'évaluation
- La Neurophysiologie Cours 2013Document17 pagesLa Neurophysiologie Cours 2013friends diaryPas encore d'évaluation
- MI3 Neurosciences Histologie Du Systeme Nerveux LavabreDocument17 pagesMI3 Neurosciences Histologie Du Systeme Nerveux LavabreDjallal HassaniPas encore d'évaluation
- Neuroanatomie Funciones-LexicoDocument69 pagesNeuroanatomie Funciones-LexicoDaniela RiosPas encore d'évaluation
- Syndrome cérébelleux_MED3_yfDocument52 pagesSyndrome cérébelleux_MED3_yfjmd9z79rbyPas encore d'évaluation
- Compression Médullaire - 3648Document10 pagesCompression Médullaire - 3648Vencesslass OndoboPas encore d'évaluation
- OK Syndrome Nerveux PériphériqueDocument7 pagesOK Syndrome Nerveux PériphériqueoslobanuandreeaPas encore d'évaluation
- Cours Mucle RénovéDocument11 pagesCours Mucle Rénovémohamed.aziz.amri.11Pas encore d'évaluation
- Cours 1Document7 pagesCours 1dchttk6htvPas encore d'évaluation
- SensibiliteDocument39 pagesSensibilitewooden latexPas encore d'évaluation
- Bases Physiopathologie de La DouleurDocument7 pagesBases Physiopathologie de La Douleurdchttk6htvPas encore d'évaluation
- UE 3 Reflexe MedullaireDocument6 pagesUE 3 Reflexe MedullaireCamille DruonPas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- Neurones Et Transmission SynaptiqueDocument6 pagesNeurones Et Transmission Synaptiquelouanne kloepferPas encore d'évaluation
- Physique Et TéléphoniqueDocument4 pagesPhysique Et TéléphoniquemeyemobiangjPas encore d'évaluation
- Anatomie 2011Document52 pagesAnatomie 2011Manal AhPas encore d'évaluation
- NRO TD8. Syndromes Médullaires (Poly)Document6 pagesNRO TD8. Syndromes Médullaires (Poly)aliabdsettarboudebzaPas encore d'évaluation
- La Moelle Épinière, Centre RéflexeDocument6 pagesLa Moelle Épinière, Centre RéflexeSaken667Pas encore d'évaluation
- Support N° 6Document9 pagesSupport N° 6Ossete BricePas encore d'évaluation
- Le Tissu NerveuxDocument10 pagesLe Tissu NerveuxANDRIANOME tafitaPas encore d'évaluation
- Voies de La Sensibilité Et DouleurDocument20 pagesVoies de La Sensibilité Et DouleurKhelfaoui NahlaPas encore d'évaluation
- Physiologie Neuro-MuscuclaireDocument53 pagesPhysiologie Neuro-MuscuclaireMampionona RakotonandrasanaPas encore d'évaluation
- Cours 8. Les Organes Des SensDocument4 pagesCours 8. Les Organes Des SensmousalsaletturkiyaPas encore d'évaluation
- L'organe Du Toucher - 100228Document8 pagesL'organe Du Toucher - 100228EsperancePas encore d'évaluation
- Résumé G.Chakib NerveuxDocument5 pagesRésumé G.Chakib NerveuxMaroua AbzPas encore d'évaluation
- Physiologie Physiopathologie Douleur PDFDocument16 pagesPhysiologie Physiopathologie Douleur PDFKAMARPas encore d'évaluation
- 1-Physiologie Des Récepteurs Sensoriels PDFDocument34 pages1-Physiologie Des Récepteurs Sensoriels PDFImad Imad ZaroutaPas encore d'évaluation
- La DouleurDocument37 pagesLa DouleurclaudedPas encore d'évaluation
- Atteintes Nerveuses RadiculairesDocument24 pagesAtteintes Nerveuses RadiculairesImane BenderazPas encore d'évaluation
- Le Syndrome PyramidalDocument5 pagesLe Syndrome Pyramidalbrahim brahimPas encore d'évaluation
- Généralités Sur La Physiologie RénaleDocument3 pagesGénéralités Sur La Physiologie RénaleZied BouabenePas encore d'évaluation
- Clairances GlomérulairesDocument1 pageClairances GlomérulairesZied BouabenePas encore d'évaluation
- Physiologie de La Glande ThyroïdeDocument5 pagesPhysiologie de La Glande ThyroïdeZied Bouabene100% (1)
- La SomesthésieDocument3 pagesLa SomesthésieZied Bouabene100% (1)
- Les Noyaux Gris CentrauxDocument5 pagesLes Noyaux Gris CentrauxZied BouabenePas encore d'évaluation
- Physiologie Des Récepteurs SensorielsDocument2 pagesPhysiologie Des Récepteurs SensorielsSaken667Pas encore d'évaluation
- Clairance Osmolaire Et Clairance de L'Eau LibreDocument1 pageClairance Osmolaire Et Clairance de L'Eau LibreZied Bouabene67% (3)
- Réabsorption Rénale Du GlucoseDocument1 pageRéabsorption Rénale Du GlucoseSaken667Pas encore d'évaluation
- Généralités Sur Le Système NerveuxDocument3 pagesGénéralités Sur Le Système NerveuxSaken667Pas encore d'évaluation
- Filtration GlomérulaireDocument3 pagesFiltration GlomérulaireZied BouabenePas encore d'évaluation
- Physiologie Du Complexe Hypothalamo-HypophysaireDocument5 pagesPhysiologie Du Complexe Hypothalamo-HypophysaireZied Bouabene100% (1)
- Mécanisme D'Acidification Des UrinesDocument3 pagesMécanisme D'Acidification Des UrinesZied Bouabene100% (1)
- Excrétion Urinaire Du Potassium PDFDocument2 pagesExcrétion Urinaire Du Potassium PDFZied BouabenePas encore d'évaluation
- Circulation RénaleDocument3 pagesCirculation RénaleZied BouabenePas encore d'évaluation
- La MictionDocument2 pagesLa MictionZied Bouabene100% (1)
- Physiologie de La SurrénaleDocument7 pagesPhysiologie de La SurrénaleSaken667100% (1)
- Réabsorption Rénale Du Calcium Et Des PhosphatesDocument2 pagesRéabsorption Rénale Du Calcium Et Des PhosphatesZied BouabenePas encore d'évaluation
- Examen NeurologiqueDocument7 pagesExamen NeurologiqueRoxana Rox100% (1)
- Physio DouleurDocument27 pagesPhysio DouleurMohamed Ali MtibaaPas encore d'évaluation
- Presentation 1Document50 pagesPresentation 1mohamed abderrazak LagariPas encore d'évaluation
- Poly Fonct 2013Document106 pagesPoly Fonct 2013Williams SantimePas encore d'évaluation
- Chapitre 1 - Le Système SomesthésiqueDocument61 pagesChapitre 1 - Le Système Somesthésiqueasma2490Pas encore d'évaluation
- Chapitre III SomesthesieDocument22 pagesChapitre III SomesthesieDenisco NkemgnePas encore d'évaluation
- 2019 Medecine GuidottiMarcoDocument125 pages2019 Medecine GuidottiMarcozoheir ouffroukhPas encore d'évaluation
- 09 Neurologie - Sémio Sandoz PDFDocument50 pages09 Neurologie - Sémio Sandoz PDFLaura PussPas encore d'évaluation
- Neuro, SomesthésieDocument1 pageNeuro, SomesthésieMarMar TsiePas encore d'évaluation
- Sensib EiaDocument37 pagesSensib Eiasylvie jacklinePas encore d'évaluation
- Les Voies de La Sensibilité OKDocument13 pagesLes Voies de La Sensibilité OKMichel S. FayePas encore d'évaluation
- Neurophysio de La FCT ProprioceptiveDocument8 pagesNeurophysio de La FCT ProprioceptiveImane BenderazPas encore d'évaluation
- 6.les Troubles de La SensibilitéDocument6 pages6.les Troubles de La SensibilitéHøpë FûllPas encore d'évaluation
- Cours de Physiologie Animale II / Module 27 / Filière SV5Document73 pagesCours de Physiologie Animale II / Module 27 / Filière SV5HamidPas encore d'évaluation
- Barat 2010Document13 pagesBarat 2010Bruno NdoumouPas encore d'évaluation