Vous aimerez peut-être aussi
- Neurobiologie: Les Grands Articles d'UniversalisD'EverandNeurobiologie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Œil humain: Les Grands Articles d'UniversalisD'EverandŒil humain: Les Grands Articles d'UniversalisPas encore d'évaluation
- 42.vestibule Membraneux Et Voies VestibulairesDocument5 pages42.vestibule Membraneux Et Voies Vestibulairesadelmiringui2Pas encore d'évaluation
- 8 - Audition Equilibration 2021Document89 pages8 - Audition Equilibration 2021Ammar ChaouiPas encore d'évaluation
- Fichier Produit 2315Document8 pagesFichier Produit 2315MaurelPas encore d'évaluation
- 11 - Audition Dr. LassakeurDocument75 pages11 - Audition Dr. LassakeurAmmar ChaouiPas encore d'évaluation
- Organes Des SensDocument12 pagesOrganes Des SensEl Zizou MerenguePas encore d'évaluation
- Histo OumaDocument7 pagesHisto OumaAnass VillaPas encore d'évaluation
- La Muqueuse Olfactive Et Les Bourgeons Du GoutDocument8 pagesLa Muqueuse Olfactive Et Les Bourgeons Du Goutjoudjojo.06Pas encore d'évaluation
- Curs 3 UrecheaDocument28 pagesCurs 3 Urecheaqty9jgkpnzPas encore d'évaluation
- COCHLEE MEMBRANEUSE Et VOIES COCHLEAIRESDocument6 pagesCOCHLEE MEMBRANEUSE Et VOIES COCHLEAIRESadelmiringui2Pas encore d'évaluation
- 23 Vestibule Membraneux Et Voies VestibulairesDocument3 pages23 Vestibule Membraneux Et Voies Vestibulairesadelmiringui2Pas encore d'évaluation
- 22 La Cochlée Membraneuse Et Voies CochléairesDocument5 pages22 La Cochlée Membraneuse Et Voies Cochléairesadelmiringui2Pas encore d'évaluation
- Anatomie Et Physiologie Du LimbeDocument4 pagesAnatomie Et Physiologie Du Limbebin ozumbaiPas encore d'évaluation
- 1 - Physiologie - Renale +++Document28 pages1 - Physiologie - Renale +++Bilel Dhaouadi100% (1)
- UE2 - Histologie - Epithéliums (Partie 2)Document11 pagesUE2 - Histologie - Epithéliums (Partie 2)sun-nee-chan9Pas encore d'évaluation
- Anatomie Et Physiologie en ORLDocument77 pagesAnatomie Et Physiologie en ORLbenben31Pas encore d'évaluation
- Anatomie IV OPHT (4. Anatomie de La Conjonctive)Document24 pagesAnatomie IV OPHT (4. Anatomie de La Conjonctive)abdo100% (1)
- Anatomie - de - Loreille - Interne 2Document16 pagesAnatomie - de - Loreille - Interne 2kadmirikawtar1995Pas encore d'évaluation
- Neuroanat-2 0Document83 pagesNeuroanat-2 0seloua kaid100% (1)
- Chapitre 11 - Les Systèmes MentalesDocument66 pagesChapitre 11 - Les Systèmes MentalesMouctarPas encore d'évaluation
- M2-Neuro - Anatomie de L'audition Et de L'équilibrationDocument12 pagesM2-Neuro - Anatomie de L'audition Et de L'équilibrationThomas HussonPas encore d'évaluation
- Anatomie de L'oeil - Systématisation Des Voies OptiquesDocument17 pagesAnatomie de L'oeil - Systématisation Des Voies OptiquesYannick NgninzekoPas encore d'évaluation
- CORDECTOMIE FDocument7 pagesCORDECTOMIE Fadelmiringui2Pas encore d'évaluation
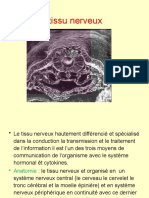
- Le Tissu NerveuxDocument51 pagesLe Tissu NerveuxYouness KhalfaouiPas encore d'évaluation
- Physiologie Animale Cours 7Document35 pagesPhysiologie Animale Cours 7Alhassane BahPas encore d'évaluation
- Anatomie-Physiologie Du Systeme Nerveux 2013041712271888Document54 pagesAnatomie-Physiologie Du Systeme Nerveux 2013041712271888Hami BenPas encore d'évaluation
- Cap Ill AiresDocument6 pagesCap Ill AiresZeroualPas encore d'évaluation
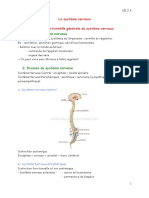
- Le Système Nerveux PDFDocument13 pagesLe Système Nerveux PDFSaggou OuafaPas encore d'évaluation
- Chapitre 3 Le Système NerveuxDocument26 pagesChapitre 3 Le Système NerveuxdfhohfeicxhPas encore d'évaluation
- Innervation Et Vascularisation de Lorgane Dentaire 150524104247 Lva1 App6891Document30 pagesInnervation Et Vascularisation de Lorgane Dentaire 150524104247 Lva1 App6891minatta0% (1)
- 7AS1Document110 pages7AS1zeinebou sidi alhadiPas encore d'évaluation
- Anatomie Des Nerfs CraniensDocument104 pagesAnatomie Des Nerfs CraniensDr Hadjer100% (3)
- 015 - Le Nerf OptiqueDocument6 pages015 - Le Nerf OptiqueGéovanixPas encore d'évaluation
- Innervation Et Vascularisation de Lorgane DentaireDocument30 pagesInnervation Et Vascularisation de Lorgane DentaireMedecine Dentaire50% (2)
- Chapitre II: Organisation Cellulaire Du Système NerveuxDocument36 pagesChapitre II: Organisation Cellulaire Du Système Nerveuxnesrine MoadPas encore d'évaluation
- Curs 2 - OchiulDocument39 pagesCurs 2 - Ochiulqty9jgkpnzPas encore d'évaluation
- Le Syste0300me NerveuxDocument7 pagesLe Syste0300me NerveuxAmel YahiaPas encore d'évaluation
- Rappel Anatomo Physiologique Du Système NerveuxDocument58 pagesRappel Anatomo Physiologique Du Système Nerveuxlina mezianPas encore d'évaluation
- Les Épithéliums (Cours)Document89 pagesLes Épithéliums (Cours)Adil AzdimousaPas encore d'évaluation
- La Moelle SpinaleDocument11 pagesLa Moelle SpinalezainabPas encore d'évaluation
- Anatomie de CCDocument12 pagesAnatomie de CCyoussef majdPas encore d'évaluation
- Complément Ronéo Anatomie N°1Document5 pagesComplément Ronéo Anatomie N°1almnaouarPas encore d'évaluation
- 4 - Histo Du Cortex CérébelleuxDocument21 pages4 - Histo Du Cortex Cérébelleuxbf ImprimeriePas encore d'évaluation
- Anatomie TOME IIIDocument32 pagesAnatomie TOME IIIHoUda ElGoPas encore d'évaluation
- Histologie de L'appareil RespiratoireDocument7 pagesHistologie de L'appareil RespiratoireAhlam BsdPas encore d'évaluation
- Oreille 2020Document36 pagesOreille 2020EbePas encore d'évaluation
- 4eme Type Tissu NerveuxDocument36 pages4eme Type Tissu Nerveuxgroupe_svi3Pas encore d'évaluation
- Le Tissu Nerveux 2022Document90 pagesLe Tissu Nerveux 2022babacar98.04Pas encore d'évaluation
- Cours VisionDocument32 pagesCours VisionSana BENALIPas encore d'évaluation
- Cours TS2 THIAWDocument117 pagesCours TS2 THIAWBaye ModouPas encore d'évaluation
- Ganglion Nerveux FinalDocument3 pagesGanglion Nerveux Finalmalakzerizer2005Pas encore d'évaluation
- Cours SVT Tle D&C 2013-2014Document17 pagesCours SVT Tle D&C 2013-2014sandwidibernard76Pas encore d'évaluation
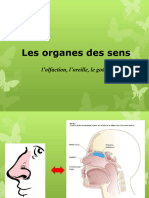
- 1 L'Audition, L'odorat, Le Goût, Le ToucherDocument24 pages1 L'Audition, L'odorat, Le Goût, Le ToucherYohanPas encore d'évaluation
- Histologie de L'oreille 2011-2012Document72 pagesHistologie de L'oreille 2011-2012Nada Affes89% (9)
- 07-Le Tissu NerveuxDocument11 pages07-Le Tissu NerveuxBemmoussat AsmaPas encore d'évaluation
- Le Système CérébelleuxDocument4 pagesLe Système CérébelleuxMohammed GartitPas encore d'évaluation
- 6-Anatomie Du Labyrinthe Membraneux 2022Document48 pages6-Anatomie Du Labyrinthe Membraneux 2022Eli AhoblePas encore d'évaluation
- Le Système NerveuxDocument88 pagesLe Système NerveuxMorad FerrahPas encore d'évaluation
- Vestibule Membraneux Et Voies VestibulairesDocument148 pagesVestibule Membraneux Et Voies Vestibulairesadelmiringui2Pas encore d'évaluation
- 15.mycoplasma (Resume)Document1 page15.mycoplasma (Resume)NA MESPas encore d'évaluation
- 1 COURS - LES - EPITHELIUMS - DE - REVETEMENT - 1 - .Docx Filename UTF-8''COURS LES EPITHELIUMS DE REVETEMENTDocument7 pages1 COURS - LES - EPITHELIUMS - DE - REVETEMENT - 1 - .Docx Filename UTF-8''COURS LES EPITHELIUMS DE REVETEMENTNA MESPas encore d'évaluation
- Examen Clinique de L'appareil DigestifDocument34 pagesExamen Clinique de L'appareil DigestifNA MESPas encore d'évaluation
- 3ème Année EMD 2 Anapath 2019 1Document8 pages3ème Année EMD 2 Anapath 2019 1NA MESPas encore d'évaluation
- Approche Syndromique de La Sémiologie Myocardique Et PéricardiqueDocument18 pagesApproche Syndromique de La Sémiologie Myocardique Et PéricardiqueNA MESPas encore d'évaluation
- DysprotéinémiesDocument38 pagesDysprotéinémiesNA MESPas encore d'évaluation
- Organe de La Gustation (Enregistré Automatiquement) PDFDocument2 pagesOrgane de La Gustation (Enregistré Automatiquement) PDFNA MESPas encore d'évaluation
- Norme FD P20-651Document1 pageNorme FD P20-651benoo13100% (1)
- Diabete SucreeDocument95 pagesDiabete SucreeRomuald ZidaPas encore d'évaluation
- Rapport de Tomate-1Document58 pagesRapport de Tomate-1Ismail Alaoui100% (6)
- BDR - FICHE - Cours 1 Mitose Et MéioseDocument15 pagesBDR - FICHE - Cours 1 Mitose Et Méiosesurgeon forgeronPas encore d'évaluation
- Anatomie Fonctionelle Rénale 2017 B DioufDocument66 pagesAnatomie Fonctionelle Rénale 2017 B Dioufexauce MondéPas encore d'évaluation
- 04 - PPT Arvalis-Rrgc - 220208 - OccDocument14 pages04 - PPT Arvalis-Rrgc - 220208 - OccMoradPas encore d'évaluation
- Genetique QCM PDFDocument5 pagesGenetique QCM PDFBouhamey Traore100% (1)
- Formation Microbiologie Cc3a9rc3a9ales Ifim 2018Document246 pagesFormation Microbiologie Cc3a9rc3a9ales Ifim 2018bessama84Pas encore d'évaluation
- Les TaninsDocument2 pagesLes TaninsYuliya RomanteevaPas encore d'évaluation
- 3AP Polycope TP S2 FMPR 2019-2020Document25 pages3AP Polycope TP S2 FMPR 2019-2020OMAR EL HAMDAOUIPas encore d'évaluation
- Liban SVT Specialite S 2019Document6 pagesLiban SVT Specialite S 2019LETUDIANTPas encore d'évaluation
- Heredité Special Medecine SMDocument7 pagesHeredité Special Medecine SMkanzaPas encore d'évaluation
- Tabla Periódica IUPACDocument1 pageTabla Periódica IUPACChristopher MorenoPas encore d'évaluation
- D. BonneauDocument5 pagesD. BonneaumitroisergiuPas encore d'évaluation
- FruitDocument36 pagesFruitAbdellahHammaPas encore d'évaluation
- La Gonarthrose Traitement Chirurgical de L' Arthroscopie A La Prothse PDFDocument725 pagesLa Gonarthrose Traitement Chirurgical de L' Arthroscopie A La Prothse PDFSergiu Plesca100% (1)
- PFEDocument68 pagesPFEMariem MannaiPas encore d'évaluation
- Chap 1 II CellulesDocument33 pagesChap 1 II CellulesOrskyfirePas encore d'évaluation
- Mots Invariables Toujours Au Pluriel - Grammaire AIDENETDocument8 pagesMots Invariables Toujours Au Pluriel - Grammaire AIDENETAssociation-adac Adac SenPas encore d'évaluation
- 5TQ Formation-Scientifique PIPERS UAA13 ExercicesDocument5 pages5TQ Formation-Scientifique PIPERS UAA13 ExercicesAudrey GustinPas encore d'évaluation
- Processus InflamatoireDocument5 pagesProcessus InflamatoireChah Ra ZedPas encore d'évaluation
- Society N8 - 12 Au 25 Juin 2015Document92 pagesSociety N8 - 12 Au 25 Juin 2015yankevPas encore d'évaluation
- Paludisme - FicheDocument4 pagesPaludisme - FicheRaja KhanPas encore d'évaluation
- TP BV Thallophytes - B233 S3 BCG-PR F MelloukiDocument21 pagesTP BV Thallophytes - B233 S3 BCG-PR F MelloukiLSTL TACQ0% (1)
- Prévalence Des Infections UrinairesDocument27 pagesPrévalence Des Infections Urinairesmaguette.mbaye08Pas encore d'évaluation
- Composition IIDocument3 pagesComposition IIkhadidja Latm100% (1)
- 010033280Document319 pages010033280RussellPas encore d'évaluation
- La BiosécuritéDocument27 pagesLa BiosécuritéKhyarhoum Brahim100% (1)
- Associations Conventionnées EN IdF 2020-2021Document1 pageAssociations Conventionnées EN IdF 2020-2021jk rPas encore d'évaluation
- TP 5Document27 pagesTP 5PierrePas encore d'évaluation