Vous aimerez peut-être aussi
- La Respiration Cellulaire 1 PDFDocument27 pagesLa Respiration Cellulaire 1 PDFyyyyy100% (1)
- Medecine FR Cours 2 Anatomie, Planches Et LéxiqueDocument216 pagesMedecine FR Cours 2 Anatomie, Planches Et Léxiqueyannick94% (32)
- Retz - Relaxer Les Enfants À L'école MaternelleDocument66 pagesRetz - Relaxer Les Enfants À L'école MaternellePatrickPas encore d'évaluation
- Toxoplasmose FinalDocument27 pagesToxoplasmose FinalAzzedine Baka100% (1)
- Document Le Texte Explicatif Conrrigé 1Document33 pagesDocument Le Texte Explicatif Conrrigé 1belkacem100% (1)
- Alternative Santé - Avril 2013Document28 pagesAlternative Santé - Avril 2013Bastien Gall de Sillé50% (2)
- S2CHP2Physiologie Cardio-Vasculaire 1-40Document40 pagesS2CHP2Physiologie Cardio-Vasculaire 1-40Noura AzaouagPas encore d'évaluation
- Fgioytr - CopieDocument462 pagesFgioytr - CopieOusmane DiakitéPas encore d'évaluation
- Liste Des Analyses Pour Le 1er Janvier 2024Document1 380 pagesListe Des Analyses Pour Le 1er Janvier 2024Khalid ChaterPas encore d'évaluation
- La Drepanocytose Probleme de L Afrique PDFDocument5 pagesLa Drepanocytose Probleme de L Afrique PDFsarahloba100Pas encore d'évaluation
- Paludisme: Classifications CIM Du PaludismeDocument33 pagesPaludisme: Classifications CIM Du PaludismeAymenSaidiPas encore d'évaluation
- NUT2006 Sa 0033Document20 pagesNUT2006 Sa 0033Kevin NoahPas encore d'évaluation
- Paludisme - Cours A6PR ALLOUNDocument21 pagesPaludisme - Cours A6PR ALLOUNPizongo RamdhinoPas encore d'évaluation
- Parasitoses Intestinales Méconnues: Objectifs PédagogiquesDocument8 pagesParasitoses Intestinales Méconnues: Objectifs PédagogiquesBest ManPas encore d'évaluation
- Paludisme Grave Enft Epee KampeDocument69 pagesPaludisme Grave Enft Epee KampeIdiAmadouPas encore d'évaluation
- 1 s2.0 S0035378704711182 MainDocument1 page1 s2.0 S0035378704711182 MainguehamsanaPas encore d'évaluation
- Épidémiologie de La Drépanocytose en France Et Dans Le Monde - La Revue Du Praticien HistoriqueDocument2 pagesÉpidémiologie de La Drépanocytose en France Et Dans Le Monde - La Revue Du Praticien HistoriqueSacire TchatcheiPas encore d'évaluation
- Paludisme Phar2 - FAPH 130224FnDocument20 pagesPaludisme Phar2 - FAPH 130224Fndoukarafousseyni7Pas encore d'évaluation
- Les Parasites Et Le Cycle ParasitaireDocument4 pagesLes Parasites Et Le Cycle Parasitaireyoussef elcoustafPas encore d'évaluation
- 5 PaludismeDocument10 pages5 PaludismeNour El HoudaPas encore d'évaluation
- DREPANOCYTOSEDocument45 pagesDREPANOCYTOSEpepePas encore d'évaluation
- Dorcas Kitoko MEMOIRE FINAL AOUTDocument51 pagesDorcas Kitoko MEMOIRE FINAL AOUTEngwataPas encore d'évaluation
- MoustiqueDocument132 pagesMoustiqueAbdel Kader JikaPas encore d'évaluation
- Travail de Recherche N°4 - Les Mutations de PlasmodiumDocument9 pagesTravail de Recherche N°4 - Les Mutations de Plasmodiumnoubaramadji.yamtiPas encore d'évaluation
- ErythroenzymopathiesDocument10 pagesErythroenzymopathiesAudrey LyliPas encore d'évaluation
- Le Paludisme 2023Document78 pagesLe Paludisme 2023gracekouassi08Pas encore d'évaluation
- 1 s2.0 S0399077X76800017 MainDocument5 pages1 s2.0 S0399077X76800017 MainO A SEIDOUNAPas encore d'évaluation
- Ankylostomose RFL2016Document4 pagesAnkylostomose RFL2016Anou KalengaPas encore d'évaluation
- 1 Paludisme L3 Med Pharm INSSADocument56 pages1 Paludisme L3 Med Pharm INSSAcheikmohamedbayoPas encore d'évaluation
- Paludisme Minimodule 2021 2022Document41 pagesPaludisme Minimodule 2021 2022AmenElleh ShilPas encore d'évaluation
- 1 s2.0 S0399077X00882852 MainDocument5 pages1 s2.0 S0399077X00882852 Mainlina aya hiourPas encore d'évaluation
- CyclosporoseDocument3 pagesCyclosporosevague2000Pas encore d'évaluation
- PaludismeDocument17 pagesPaludismeiri NaPas encore d'évaluation
- Pec PaluDocument74 pagesPec PaluIdiAmadouPas encore d'évaluation
- Cours Thème 3A - Chapitre 1 Variation Génétique Et SantéDocument10 pagesCours Thème 3A - Chapitre 1 Variation Génétique Et Santébtbines2007Pas encore d'évaluation
- Goyffon 3Document8 pagesGoyffon 3yaya1310Pas encore d'évaluation
- Biosys Gly 2Document8 pagesBiosys Gly 2Dina BougandouraPas encore d'évaluation
- Hellp Syndrome Revelant Une Necrose à Â'sophagienne :a Propos Dun CasDocument5 pagesHellp Syndrome Revelant Une Necrose à Â'sophagienne :a Propos Dun CasIJAR JOURNALPas encore d'évaluation
- La NutrigénétiqueDocument3 pagesLa NutrigénétiqueAmina BelaidPas encore d'évaluation
- CAPM IT CAT Chardon A GluDocument3 pagesCAPM IT CAT Chardon A GluBoumadian SviPas encore d'évaluation
- Predavanje 02 Protozoa 2Document29 pagesPredavanje 02 Protozoa 2VanjaSekulicPas encore d'évaluation
- HENRY - Benoit Splénomégalie, Déformabilité Des Globules Rouges Circulants PlasmodiumDocument261 pagesHENRY - Benoit Splénomégalie, Déformabilité Des Globules Rouges Circulants PlasmodiumSalam SawadogoPas encore d'évaluation
- Cours Path Asc 1Document59 pagesCours Path Asc 1Ouattara BamoryPas encore d'évaluation
- Diagnostico y EpidemiologiaDocument9 pagesDiagnostico y EpidemiologiaWalter Huamani AlvitesPas encore d'évaluation
- 434 1591 1 PB PDFDocument7 pages434 1591 1 PB PDFMourad MhPas encore d'évaluation
- Ann. Soc. Belge Méd. Trop.: A. FainDocument9 pagesAnn. Soc. Belge Méd. Trop.: A. FainWillson Peter Dungu businessPas encore d'évaluation
- 393-400 COM AFFICHEES Session ParasitologieDocument8 pages393-400 COM AFFICHEES Session ParasitologieAnou KalengaPas encore d'évaluation
- Système DuffyDocument5 pagesSystème Duffydhikra abdelPas encore d'évaluation
- Déficit en G6PD, en Pyruvate Kinase, en Pyrimidine 5' NucléotidaseDocument1 pageDéficit en G6PD, en Pyruvate Kinase, en Pyrimidine 5' NucléotidaseHarouna RachidPas encore d'évaluation
- EnzymopathiesokDocument8 pagesEnzymopathiesoksaydou malgoubriPas encore d'évaluation
- Horizons Hemato Mars 2014-35-36 PissardDocument3 pagesHorizons Hemato Mars 2014-35-36 PissardInasse MourabitiPas encore d'évaluation
- PALUDISMEDocument4 pagesPALUDISMEiranithia96Pas encore d'évaluation
- Neurotoxicité Des Substances NaturellesDocument8 pagesNeurotoxicité Des Substances NaturellesaissaPas encore d'évaluation
- DrépanocytoseDocument11 pagesDrépanocytoseWilfried AboPas encore d'évaluation
- Ascaris LumbricoidesDocument16 pagesAscaris LumbricoidesalmnaouarPas encore d'évaluation
- g6pd 2Document7 pagesg6pd 2pmat16Pas encore d'évaluation
- J Tracli 2014 08 139Document4 pagesJ Tracli 2014 08 139noe essonokolloPas encore d'évaluation
- RP 195 Moreno-SabaterDocument6 pagesRP 195 Moreno-Sabateralkaousissoko71Pas encore d'évaluation
- Anémies Dysérythropoïétiques Congénitales PDFDocument4 pagesAnémies Dysérythropoïétiques Congénitales PDFzlimitounePas encore d'évaluation
- CERTAD2015Peixes Suiça BomDocument13 pagesCERTAD2015Peixes Suiça BomBrayan KuraharaPas encore d'évaluation
- RisquesparasitairesDocument1 pageRisquesparasitairesfelix.tiako-kameniPas encore d'évaluation
- Paludisme Grave Mangistrale 2023 2023Document43 pagesPaludisme Grave Mangistrale 2023 2023Ange RaphaëlPas encore d'évaluation
- Pnds Hypoparathyroidie VFDocument35 pagesPnds Hypoparathyroidie VFmalabarPas encore d'évaluation
- Biomolécule Alcaloïdique (Quinine Ou Chloroquine)Document24 pagesBiomolécule Alcaloïdique (Quinine Ou Chloroquine)Riadh ChouaibiPas encore d'évaluation
- Le cancer - Maladie de la civilisation? (Traduit): Une étude anthropologique et historiqueD'EverandLe cancer - Maladie de la civilisation? (Traduit): Une étude anthropologique et historiquePas encore d'évaluation
- Devoir UCAD 2019 3e AnDocument12 pagesDevoir UCAD 2019 3e AnMbaye NDOYEPas encore d'évaluation
- Devoir EUROMED L3S6 2019Document3 pagesDevoir EUROMED L3S6 2019Mbaye NDOYEPas encore d'évaluation
- Page de GardeDocument1 pagePage de GardeMbaye NDOYEPas encore d'évaluation
- A L'attention Du Chef de Département de Génie Civil: Fenêtre Mermoz Cité Des Enseignants Du SupérieurDocument1 pageA L'attention Du Chef de Département de Génie Civil: Fenêtre Mermoz Cité Des Enseignants Du SupérieurMbaye NDOYEPas encore d'évaluation
- Methode de Gauss SeidelDocument2 pagesMethode de Gauss SeidelMbaye NDOYEPas encore d'évaluation
- Demande D'emploi - SignéDocument1 pageDemande D'emploi - SignéMbaye NDOYEPas encore d'évaluation
- DEVOIRDocument22 pagesDEVOIRMbaye NDOYEPas encore d'évaluation
- CM - Eléments D'ingerie Des Fondations 1Document44 pagesCM - Eléments D'ingerie Des Fondations 1Mbaye NDOYEPas encore d'évaluation
- Wa0022.Document3 pagesWa0022.Mbaye NDOYEPas encore d'évaluation
- DEVOIR DDocument9 pagesDEVOIR DMbaye NDOYEPas encore d'évaluation
- Conduction Corrigés TDDocument8 pagesConduction Corrigés TDMbaye NDOYEPas encore d'évaluation
- Note de Calcul Pont TangerDocument13 pagesNote de Calcul Pont TangerMbaye NDOYEPas encore d'évaluation
- Chap4 Premier Principe ThermodynamiqueDocument28 pagesChap4 Premier Principe ThermodynamiqueMbaye NDOYEPas encore d'évaluation
- Chap5.1 ThermoDocument21 pagesChap5.1 ThermoMbaye NDOYEPas encore d'évaluation
- Correction ConvectionDocument8 pagesCorrection ConvectionMbaye NDOYEPas encore d'évaluation
- Engins D'exécutions Des Chaussées Souples Et Des Chaussées RigidesDocument16 pagesEngins D'exécutions Des Chaussées Souples Et Des Chaussées RigidesMbaye NDOYEPas encore d'évaluation
- Exercice RDMDocument2 pagesExercice RDMMbaye NDOYEPas encore d'évaluation
- 2.1 FP Radio-Anatomie Hépatique UtileDocument163 pages2.1 FP Radio-Anatomie Hépatique UtileDiana CsillaPas encore d'évaluation
- OsteopontinDocument2 pagesOsteopontinLilia KourdachePas encore d'évaluation
- Le Squelette Et Les Os - AlloprofDocument12 pagesLe Squelette Et Les Os - AlloprofJoëlPas encore d'évaluation
- These El Bachaoui Nadim UirDocument12 pagesThese El Bachaoui Nadim UirNadim el bachaouiPas encore d'évaluation
- CONCOURS Biologie 2016Document3 pagesCONCOURS Biologie 2016Evariste Galois NshimirimanaPas encore d'évaluation
- Anatomie IIDocument39 pagesAnatomie IIran saPas encore d'évaluation
- Cours 1: Introduction: Médecine Physique Et Réadaptation FonctionnelleDocument24 pagesCours 1: Introduction: Médecine Physique Et Réadaptation FonctionnelleJorianeC.MbembaPas encore d'évaluation
- 2016 Corps Humain PDFDocument47 pages2016 Corps Humain PDFZoulkiffiPas encore d'évaluation
- AngorDocument26 pagesAngorAngelaPas encore d'évaluation
- Exposé Sur Le Pigeon G9Document24 pagesExposé Sur Le Pigeon G9dembele9242Pas encore d'évaluation
- Correction Fiche TP1 A3C1ST1T1 2ndDocument4 pagesCorrection Fiche TP1 A3C1ST1T1 2ndChahlaa0% (1)
- DavenportDocument28 pagesDavenportCheick SANOUPas encore d'évaluation
- TD Structure Des GlucidesDocument8 pagesTD Structure Des GlucidesSomeone IdkPas encore d'évaluation
- CatecholamineDocument29 pagesCatecholamineErick AndersonPas encore d'évaluation
- Concours Blanc 6 BiologieDocument8 pagesConcours Blanc 6 Biologiejunior gatientPas encore d'évaluation
- Embryologie Des Os Du Crane Et de La FaceDocument40 pagesEmbryologie Des Os Du Crane Et de La FaceDavid BienvenuePas encore d'évaluation
- Alchimie The WitcherDocument2 pagesAlchimie The Witcherluno dsPas encore d'évaluation
- Néphrectomie Simple Et Néphrectomie Elargie Pour CancerDocument28 pagesNéphrectomie Simple Et Néphrectomie Elargie Pour Cancerdjasmi mohamedPas encore d'évaluation
- Physiologie de L'appareil Respiratoire - PR FaroudyDocument14 pagesPhysiologie de L'appareil Respiratoire - PR FaroudyMarouane bouhabsPas encore d'évaluation
- II-1 Introduction: Chapitre II Description Et Fonctionnement de L'automateDocument25 pagesII-1 Introduction: Chapitre II Description Et Fonctionnement de L'automateDan BiPas encore d'évaluation
- Biology Paper 2 TZ2 HL MarkschemeDocument19 pagesBiology Paper 2 TZ2 HL Markschemejosemariaperez-carasa2006Pas encore d'évaluation
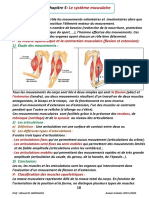
- Chapitre 5:: Le Système MusculaireDocument5 pagesChapitre 5:: Le Système MusculaireRAYANE YTPas encore d'évaluation
- 2 Cours ChronobiologieDocument3 pages2 Cours Chronobiologiecamille-puechPas encore d'évaluation
- Cours Chapitre IDocument12 pagesCours Chapitre IHou TrtyPas encore d'évaluation