Académique Documents
Professionnel Documents
Culture Documents
Bactériologie Fondamentale
Transféré par
coursenligne.lille1Titre original
Copyright
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentBactériologie Fondamentale
Transféré par
coursenligne.lille1Microbiologie 2
Chapitre 2 : La bactrie.
I. Composition chimique. 1. Lorganisme. Escherichia coli : On rcupre une quantit de bactries de lordre de 100 mg (par filtration ou par centrifugation). Les 100 mg forment une masse compacte, une pte. On dessche cette pte et on dcouvre quil y avait 70% deau. Maintenant que lon connat la masse sche, on peut peser les diffrents lments. Carbone Oxygne Azote Hydrogne Phosphore Potassium Soufre Calcium Magnsium Chlore Fer Autres % poids sec 50 20 14 8 3 2 1 0.5 0.5 0.5 0.2 0.3 Cumul 50 70 84 92 95 97 98 98.5 99 99.5 99.7 100
Lanalyse organique dune espce bactrienne Dpend de conditions de croissance et de ltat de croissance. Page 8 poly. Toutes les bactries on en commun un plan dorganisation cellulaire qui leur est particulier = procaryote. Les procaryotes nont pas e noyau diffrenci (= pas de membrane nuclaire). Les bactries taient analyses sur le plan biochimique, absence ou prsence de x. Le problme des procaryotes est quil ny a pas de reproduction sexue. La notion est donc beaucoup plus difficile mettre en place. On arrive tout de mme mettre en place des espces. Espces = grand nombre disolats naturels qui vont prsenter des caractristiques ou caractres identiques. Isolats = souches. Un trs grand nombre disolats = espce. Lespce est : - une donne artificielle cre par lHomme vrai et faux. - donne trs judicieuse et valide qui a une relle signification physiologique et naturelle. Des virus vont venir attaquer les bactries = bactriophages.
Microbiologie 2 La majorit des mmes isolats auront les mmes types de phage. Malgr le continuum volutif, la notion despce est tout de mme valable chez les bactries. La notion de souche pose problme. Souche = toues les bactries dune mme espce qui possdent quelques diffrences. Mais les mutants crer en gntique sont aussi appels mutants. Il existe donc 2 dfinitions diffrentes : - isolats naturels dune mme espce avec quelques diffrences. - rsultat du travail des chercheurs qui ont cr des mutants. En gnral, on donne des numros de souches lorsque ce sont des mutants. Lvolution est continue. Les cultures prolonges provoquent des mutations que lon ne souhaitait pas. Prolong = changer de milieu tous les 15j par exemple. Il faut trouver un mode de conservation des bactries qui limite la croissance de bactries pour limiter les mutations. 2 mthodes utilises : - lyophilisation des souches. - placer les souches basse temprature (-80C) avec un cryoprotecteur. 2. Le milieu. Les bactries sont unicellulaires et bien trop petites pour contrler le milieu extrieur. Tout un ensemble de mcanismes vont leur permettre dajuster leurs constituants cellulaires en fonction du milieu extrieur (pH, osmolarit). Ces mcanismes vont leur permettre de percevoir le milieu extrieur et sont indispensables la vie. Pour une mme bactrie, dans 2 milieux diffrents, il y aura des protines diffrentes synthtises. 3. Ltat de croissance. Si on place les bactries dans un nouveau milieu, il va falloir un certains temps pour que toutes les bactries soient dans le mme tat. Toutes les bactries vont devoir faire un ajustement de leurs constituants cellulaires. La dure va tre variable car : - dpend de la diffrence des 2 milieux (do la bactrie vient et ou elle est installe) - dpend de ltat individuel de la bactrie (une bactrie peut sadapter + vite quune autre). Aprs cet ajustement, on entre en phase exponentielle. Une fois dans ces conditions, les bactries vont toutes rester dans le mme tat pendant un certains temps, et cet tat va rester tant que le milieu nest pas modifier par la croissance = tant que les nutriments seront en quantits saturantes. On dit, dans ce cas, que la croissance est balance car on peut tre assur pendant ce temps l que tous les composants de chaque cellule augmentent par mme unit de temps. Cest la situation idale pour tudier les bactries. 4. La dynamique. 96% des cellules sont composes de macromolcules. La bactrie nest pas trs diffrente dune cellule eucaryote part 2 constituants : la mureine et les lipopolysaccharides. On ajoute au milieu du glucose radioactif marqu uniformment au carbone 14. Ensuite, on prlve et on fait une analyse des diffrents constituants et a donne : 2
Microbiologie 2 Voir schma 7 Le flux de glucose lintrieur de la cellule est trs rapide et la transformation en petites molcules est trs rapide aussi. Par contre, le flux est beaucoup + lent pour les macromolcules. Dans ce cas, au bout dun doublement, on na que la moiti. Les ARNm dmarrent + lentement que les petites molcules mais bien que ce soit des macromolcules, le temps est + rapide tout de mme. La rapidit des ARNm est particulirement cruciale chez les bactries. II. Taille et composition dune cellule moyenne. 1. Isolement et dnombrement des bactries. On va vouloir sparer les bactries les unes des autres et on va les compter. Pour cela, on va avoir plusieurs mthodes. Chaque mthode a ses avantages et ses inconvnients. Le dnombrement des B par talement. Cette mthode est utilise pour isoler une souche. Schma 8. Si lon ne connait pas bien la bactrie, on tale toutes les solutions de 10-1 10-7. Au dpart, il y a sur la boite un tapis bactrien. Puis petit petit, au fur et mesure des dilutions, on voit apparaitre des colonies. Au dpart, on des colonies qui se touchent mais elles sont toujours incomptables. Puis, on obtient des colonies isoles avec de lespace entre elles et dans ce cas, on peut facilement compter le nombre de colonies. Il faut au moins avoir 100 colonies par boite car lorsque lon va prlever le milieu, on nest pas dans une solution mais dans uns suspension cd que mme si lon ne les voit pas, il y a des bactries. Donc, quand on prend du milieu avec la pipette, on va prendre des bactries avec. A la fin, il y a donc une dviation standard. Ex : on compte 100 colonies 100 + ou 10 (10%) (Racine carr) 625 + ou 25 (4%) Il va falloir faire un compromis entre une dviation standard et lerreur de comptage de colonies. On peut galement compter la boite de dilution au dessus et celle en dessous pour vrifier si le travail a bien t fait. Dnombrement total. Schma 9 On rempli le quadrillage avec les bactries : 10-5 L. Au microscope, on va pouvoir dnombrer le nombre de bactries. Ici, on compte toutes les bactries mortes ou vivantes. Et on ramne systmatiquement en nombre de bactries / ml de culture de dpart.
Microbiologie 2 Turbidimtrie. On visualise laugmentation du trouble de la suspension bactrienne. Lappareil utilis est le spectrophotomtre ou le nphlomtre. Dans le cas du spectrophotomtre, on a : Le rcepteur mesure la lumire reue. Dans ce cas la loi de Beer Lambert ne sapplique quaux solutions et ici on a une suspension. Donc on ne mesure pas une absorbance ici. On mesure une quantit de lumire qui va tre, pendant un tps, proportionnelle au nombre de bactries. Plus la concentration de bactrie est leve, plus la lumire difracte est importante. Cest valable jusquune certaine valeur, partir dune densit de bactries trop importante ce nest plu proportionnel. Donc dans les cas qui ns intresse, on a une linarit entre une DO de 0,1 et 0,5. Schma 11 A partir du moment o on est DO = 0,5, il faudrait diluer les cultures pour toujours se trouver dans une bonne DO pour que la courbe soit toujours linaire. La DO ici ne nous a pas donn un nombre de bactrie. Le nombre de bactrie peut tre dtermin de 2 manires : - relation entre DO et poids sec des bactries - dnombrement : une DO particulire on prlve un chantillon, on dilue et on tale sur boite et aprs on compte. Il faut absolument faire une courbe dtalonnage ds quon change de spectrophotomtre. Puisquon mesure laugmentation dun trouble, ce nest pas universel pour toutes les bactries. Si on a une bactrie plus grosse, il ny aura pas le mme nombre de bactrie pour une mme DO. 2. Taille. Schma 12. Pour E. coli : Dans 100mg, on a 1011 bactries. Masse dune cellule ici = 9,5.10-13 g. Lorsquon sche cette bactrie, masse = 2,8.10-13g Taille extrmement faible. Toute petite cellule. Cette taille est trs importante par rapport la manire de vivre. Grce cette petite taille, le rapport surface volume est important. Plus il y a possibilit dchange rapide avec le milieu extrieur. Entre trs rapide de nutriment et sortie trs rapide des dchets. Permet taux mtabolique trs lev participe la croissance rapide de la majorit des bactries. Le taux mtabolique dune bactrie est de 10 100 fois suprieur que celui dune cellule eucaryote. 3. Varits et nombre des molcules. 4. Les protines. On peut faire une lectrophorse en 2 dimensions. 4
Microbiologie 2 Aprs analyse, on peut montrer quon va avoir peu prs dans une bactrie donne : 1 800 protines. Le poids moyen dune protine est de 40 000. Autrement dit : il y a environ 2,4 millions de bactries en tout dans la cellule bactrienne. La nature des 1 800 protines dpend trs troitement du milieu de culture. 5. Les ARN. Il y a 20% dARN dont 81% dARNribosomique On peut dnombrer 18 700 copies de chacune des sortes dARNr (23S, 16S, 5S) 18 700 ribosomes. 15% dARNt pour E. coli il y en a 200 000 copies. Chacun des ARNt nest pas en nombre identique, il y a environ 60 ARNt diffrents. Il reste donc 4% dARNm, ils st trs instables, la dure moyenne de demi vie dun ARNm est de lordre d1 min. Instabilit car va de 0,5 min 10min. On peut considrer, qu un moment donn, on a 600 900 diffrentes espces dARNm. On a 1 800 protines cela signifie quune partie des ARNm est organise en oprons. Opron = normalement un ARNm code une protine. Dans le cas dun opron, il y a plusieurs protines codes par le mm ARNm cd quil y a un mme promoteur pour une 2 ou 3 protine. Schma 3. Les 3 gnes ABC vont tre transcrits par le mme promoteur et donns 3 protines diffrentes qui vont tre traduites indpendamment. Le renouvellement (= turn over) est trs important. Leur instabilit participe une croissance rapide et une adaptation rapide du milieu. 6. LADN : chromosome et plasmides. LADN procaryote na pas vraiment la mme structure quun chromosome. Cette seule structure circulaire sappelle le gnome. Structure accessoire = plasmide = partie variable accessoire du gnome. Seule la molcule circulaire va tre indispensable la vie. On a une molcule circulaire double brin dont la longueur chez E. coli = 4 640 000 pb. (Pas norme). Si on prend cette molcule et quon ltire, elle fait 1 mm de long. La molcule dADN est donc fortement compacte grce des protines (qui ne sont pas des histones). Potentiellement, la protine moyenne = 40 000. Chez E. coli, la capacit maximale de codage est de 4 300 gnes donc = environ 4 300 protines. Mais, il y a une centaine de gnes. Et il y a des parties non codantes entre les gnes. On dit donc que chez E. coli, il y a environ 4 200 protines. Dans une condition donne, on a 40% 50% du gnome qui est en activit. 7. Les lipides. Les lipides reprsentent 9%. Tous les lipides sont membranaires. Pas de rserve en lipide (seul rserve = glycogne). Tous les lipides sont des phospholipides (sauf le LA de la Gram-). Schma 14 5
Microbiologie 2 X est la partie hydrophile qui peut tre variable selon le milieu et selon les bactries. 8. Les lipopolysaccharides. Lipopolysaccharide : 3,5 % Nest prsent que chez Gram-. Elle est variable selon les espces voir selon les souches. Cest une structure hautement antignique. Rle de protger la bactrie. 9. La murine. Murine : 2,5 % de la masse sche. Elle se trouve dans le peptidoglycane. Absent dans une toute petite quantit de bactries. Mais dans la grande majorit, il y a de la murine. La murine va former une coque rigide tout autour de la bactrie qui permet de se protger de la pression osmotique et donne la forme la bactrie. 10. Les molcules organiques. Voir poly page 10 table 2 Il y a des acides amins, Nuclotides et des sucres et leurs prcurseurs. 11. Les ions organiques. Voir poly page 10 table 3 Il y a des ions. Le sodium nest pas indispensable. Il est indispensable seulement pour quelques bactries (marines ou photosynthtiques). Ceci nous donne une ide de la complexit de la cellule bactrienne. La cellule bactrienne est complexe, sa composition va tre rgle de manire trs stricte mais a une adaptation trs rapide. Mais finalement la cellule bactrienne nest pas fondamentalement diffrente des autres cellules. III.Organisation de la cellule procaryote. Cest dans les annes 50 que lon a vu pour la 1re fois cette structure grce lamlioration du microscope. Permet de voir lultra structure des bactries. Lorganisation globale de la cellule est trs diffrente de celle que lon observe chez les eucaryotes. Point commun : ces cellules sont entoures dune membrane. Il ny a pas de rseaux membranaires lintrieur de la bactrie. Une seule structure membranaire. Pas dorganites lintrieur des bactries (pas de chloroplastes..). On voit un cytoplasme granuleux et qui traduit lextrme richesse en ribosomes. On voit une masse = ADN qui est dans le cytoplasme. Pas de membrane nuclaire donc pas de noyau. On a, la place, un nuclode. Labsence de membrane nuclaire fait que le chromosome baignant dans le cytoplasme, lARNm va tre en contact avec les ribosomes et donc transcription et traduction se font en mme temps. Permet une rapidit dadaptation suprieure, permet de gagner du temps pour la
Microbiologie 2 croissance balance. Mais une grande partie des avances observes chez les eucaroytes ne sont pas observables chez les bactries. Dans certains cas, il y a des structures annexes : - les fibrilles (pili) : pour adhrer en gnrale - les flagelles : pour se dplacer Voir poly page 11 et 12. Elle est petite. Non compartimente par des membranes entre ADN et machinerie. Pas dorganites complexes. Le transport des e- a lieu la surface des membranes cellulaires. On a des mcanismes de transports trs dvelopps qui vont permettre Le matriel gntique, division cellulaire et sont de complexit minimale par rapport aux cellules eucaryotes. Le flagelle est de constituions beaucoup + simple que chez les bactries. 1. Varits dans la structure de lenveloppe. Cette varit a t mise en vidence pour la 1re fois par le test de Gram. Gram = microbiologiste danois en 1884. Ce test permet de distinguer les bactries gram - et Gram+. Il faut taler les bactries sur une lame, fixer ces bactries par la chaleur. Colorer au violet de gentiane. On applique une solution diode qui va former un complexe avec le violet de gentiane. On voit que le complexe peut ressortir de certaines bactries. Si le complexe ressort de certaines bactries : elles sont appeles Gram-. Si le solvant polaire ne peut pas ressortir de la bactrie : elles sont appeles Gram+. Permet de montrer les diffrences dans la composition de lenveloppe qui ont t compris que 80 ans + tard. Cette coloration Gram dpend des conditions physiologiques. Dans certaines conditions, des Gram+ ne sont pas colores. Certaines bactries chappent cette coloration car elles ne possdent pas de paroi et donc pas murine. 2. Arches. Un grand groupe de procaryote : les arches = 2me grand groupe (aprs les bactries) de procaryote qui eux nont pas de murine et ne sont donc pas sensibles la coloration. Ces arches sont des procaryotes galement. La structure des lipides est trs diffrente. Ils ont des caractristiques communes avec les bactries et les eucaryotes. Au dbut, on pensait que ctait des bactries spciales qui vivaient dans des milieux extrmes. 3. Les 3 domaines : Archea, Bacteria, Eucarya. Voir poly page 13.
Microbiologie 2
Chapitre 3 : Lenveloppe Bactrienne.
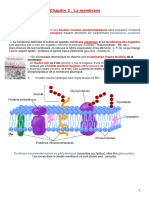
Les proprits structurales et fonctionnelles de lenveloppe vont avoir des diffrences dans leur capacit adaptative. Les diffrents constituants de lenveloppe bactrienne vont tre adapts en 1 er lieu la de nutriments. Lentre des nutriments ne doit pas se faire au dtriment de la bactrie, il faut que les produits nocifs restent lextrieur. Enveloppe : proprit dadhrence et adapte au transfert gntique. Dans le gnome bactrien des gnes sont impliqus dans la synthse, la rgulation et la maintenance des constituants de lenveloppe. Parmi les constituants de lenveloppe, on trouve des caractristiques biochimiques trs spcifiques qui permettent de classer les bactries dun point de vu taxonomique. La constitution de lenveloppe des bactries est spcifique dune espce voir dune souche. Le peptidoglycane est coll la membrane externe chez les bactries Gram - grce une lipoprotine. Il y a une + grosse couche lextrieur chez les Gram+ que chez les Gram-. Schma 1 LPS participe la protection des bactries Gram- face au milieu extrieur. I. La membrane cellulaire. 1. Les lipides. Dans cette membrane cellulaire, on trouve des lipides et des protines. Bicouche lipidique. Chez la quasi totalit des bactries, la bicouche nest forme que de phospholipides (pas de glycrol). 2 espces sont majoritaires : - les phosphatidyl glycrol (20%) - les phosphatidyl thanolamine (75%) - cardiolipide (5% minoritaire) Poly page 10 fig 5 Les 2 phosphatidyl reprsentent la tte polaire. La nature de lacide gras dtermine la + ou grande fluidit de la membrane. La membrane doit tre suffisamment fluide pour que tous les constituants puissent bouger lintrieur. Schma 2 Ex : Pourcentage dacide gras quil y a dans les phospholipides : 37% E. coli 43% acide palmitique C16 : 0 (=Satur) 33% acide palmitolique C16 : 1 (= insatur 1 fois) 23% acide cis- vaccnique C18 : 1 (= insatur 1 fois)
Microbiologie 2 1% de traces On observe au maximum 1 insaturation. 2. Les protines. Chez E. coli, 70% de la masse de la membrane reprsente des protines. Cest beaucoup + que dans la majorit des cellules eucaryotes. Beaucoup + car dans la membrane cellulaire des bactries, il y a un nombre de fonctions beaucoup + lev que dans celle des eucaryotes. Fonctions : - barrire osmotique - transport spcifique de soluts (nutriments et ions) - toutes les protines ncessaires la synthse des lipides - en grande partie les protines ncessaires la biosynthse du peptidoglycane - tous les systmes de scrtion qui vont permettre de traverser la membrane cellulaire et galement de traverser la membrane externe (Gram-) pour exporter des protines dans le milieu extrieur. - chaine respiratoire - sgrgation des chromosomes - Chimiotactisme = toutes les protines qui permettent la bactrie de percevoir des variations et les transmettre au flagelle - protines qui sont des capteurs et qui peroivent tout type de variation et qui vont transmettre lintrieur de la cellule des intgrateurs. II. Les couches externes de lenveloppe. 1. La paroi des bactries Gram positif. La rgulation de la quantit des protines dans lenveloppe est trs stricte et trs prcise. Donc un grand nombre de gne est impliqu dans la rgulation de ce nombre de protines. Les bactries vivent souvent dans un milieu hypotonique dont : Une membrane cellulaire seule ne permet pas de rsister la pression cest pourquoi, on trouve au dessus des couches externes qui permettent la bactrie de rsister des variations de pression trs importantes. Peptidoglycane : Mme les bactries Gram- (qui ont une petite couche de peptidoglycane) sont capables de rsister une pression de 100 atm. Gram+ : paisse couche de peptidoglycane permet de donner sa forme la bactrie. Les 2 acides sont lis au peptidoglycane ce qui permet de rester attacher la cellule. La structure du peptidoglycane est globalement la mme pour les Gram+ et les Gram-. Le PG (= peptidoglycane) est constitu de : - acide N_actyl_muramique (NAM) - N_actylglucosamine. (NAG) Ces 2 l rpts n fois. Ce nest pas une glycoprotine, cest un tetrapeptide (muramique). 2
Microbiologie 2 Page 17 a Un pontage permet dassocier entre elles les diffrentes chaines. Page 17 b Les 4 AA lis la NAM sont des AA particuliers. La liaison entre les 2 chaines se fait entre D alanine et la mesodiaminopimelate Page 18 a Contrairement avant, les 2 chaine peptidiques ne sont pas relies directement mais relies par des glycines. Mais leffet est le mme. Seule chose invariable = enchainement des sucres Choses variables = AA + type de pontage Mais la structure globale va tre a peu prs la mme. Page 18b A. Les pontages sont extrmement importants. B. Il y a un pontage de temps en temps. Seul 25 % des ttrapeptides sont impliqus dans les pontages. Diffrence entre ces bactries au niveau osmotique. Bactrie Gram- : Autre structure enchsse dans PG : acide tichoque et important pour lattachement la cellule. Ce sont des dterminants antigniques reconnus par lhte quand la bactrie passe dans un eucaryote. 2 grandes catgories dacides tichoques : - fabriqu par un enchainement de ribitol - fabriqu par un enchainement de molcules de glycrol Dans ces enchainements, il y a aussi des substitutions par des AA. Peptidoglycane + acide tchoque donnent une structure extrmement hydrophile obstacle pour les molcules hydrophobes cd que leur accession la membrane cellulaire va tre beaucoup + lente. Protection contre explosion de la cellule et protection contre les molcules toxiques et hydrophobes. Le peptidoglycane nest pas un obstacle pour lADN, ni pour les Place dans un milieu hypotonique, une bactrie forme un lysozyme dtruit et dgrade le peptidoglycane. Lorsque le peptidoglycane est dgrad condition dtre dans milieu hypotonique on observe immdiatement la lyse de toutes les cellules. Mme exprience dans milieu lgrement hypertonique pas de lyse mais les cellules sont dformes, on observe des protoplastes. Puis, si on remet dans milieu normal, la bactrie redevient normale. 2. La murine.
Microbiologie 2 3. La paroi et la membrane externe des bactries Gram ngatif. Pour les Gram- : On a un peptidoglycane + membrane externe. La membrane externe remplace la grosse couche de peptidoglycane de la Gram+. La membrane externe a une structure en bicouche lipidique qui est compltement asymtrique (contrairement la membrane cellulaire), la face interne de cette membrane est majoritairement constitue de phospholipides, et sur la face externe on a que la partie lipidique du lipopolysaccharide. Permet de rsister des nombreux composs toxiques. 4. Le lipopolysaccharide. Le lipopolysaccharide est compos 3 parties : - Lipide A : constitu comme un lipide : - partie hydrophile = 2 molcules de glucosamine relies 4-6 acide gras (acide gras de C12 C16 dans la majorit des cas saturs + rigide). Les glucosamines sont en gnrale phosphoryls donc chargs. Du coup, pour que les molcules du LPS interagissent ensemble pour former une barrire, il faut neutraliser ces charges avec des molcules de Ca 2+ ou Mg2+. Si la charge nest pas neutralise peut avoir des perturbations dans la structure de la membrane. - partie hydrophobe Page 19 KDO, heptose : sucres particuliers qui constituent la 1re partie de la partie polysaccharidique. Le KDO = acide cto desoxyoctanoque (pas savoir ce que veut dire KDO). KDO et hexose sont relativement bien conservs chez toutes les espces. Au dessus du noyau est greff lantigne O indispensable pour la virulence des bactries. Parmi les (4 sucres)n, il peut y avoir beaucoup denchainements (45). Page 20 Il y a des protines donc chargs structure hydrophile. Page 21b Structure que certains pensent du LPS. Certains pensent que si la partie polysacharidique est recourbe meilleure protection. La membrane externe + LPS : protection contre les molcules hydrophiles et les molcules hydrophobes. Cette structure est plus efficace pour empcher les molcules de rentrer que la protection des Gram+. Chez Gram-, seul le assure la fonction de barrire. EDTA : tous les ions bivalents sont Dsolidarise les molcules de LPS : cre des trous temporaires dans le LPS permet au lysozyme de rentrer et donc de dtruire le peptidoglycane et donc la lyse de la bactrie dans le milieu hypotonique. On obtient des sphroplastes (pas protoplaste car 2 membranes). Des rsidus de membrane externe restent attachs la membrane interne.
Microbiologie 2 LPS a un rle important dans la pathognse. Lipide A = partie la + ractive du LPS. Si trop de lipide A peut provoquer la mort. Aucune molcule nest donc sense rentrer par cette membrane externe. Ce sont des protines qui permettent le passage de nutriments. Les protines effectuent le mme travail que les protines de transport de la membrane cellulaire. On va avoir 2 catgories de protines qui permettent le transport de molcules hydrophiles : - porines : chez E. coli il y en a 2 : OmpC et OmpF, ce sont des trimres. Chaque monomre possde son canal de diffusion. Transport non spcifique. Permet de transporter des molcules hydrophiles de masse molculaire infrieure 700Da = majorit des nutriments. Ces 2 protines (Omp..) font parties des protines majeures de la membrane externe chez E. coli. - transporteurs spcifiques : permet de faire entrer des vitamines (vit B12), les phrichromes (ortho) (FhuA), des oligosaccharides (de 7-10 sucres) = petit produit de dgradation de lamidon. - Protines de structure : - OmpA : protine transmembranaire compltement dvolu la structuration de lenveloppe. - lipoprotine de Braun : permet de retenir le peptidoglycane contre la membrane externe. Possde 3 acides gras. Si absente, la bactrie ne survie pas. Donc indispensable. - Des protines impliques dans la perception du milieu extrieur. Pour expliquer les liaisons : On peut imaginer des connexions entre membrane cellulaire et membrane externe. On aurait des zones de Bayer. Schma 3. On ne connait pas trop la nature des jonctions entre membrane cellulaire et membrane externe. III.Le priplasme. Le priplasme forme un gel dans lequel on retrouve : - le peptidoglycane - diffrentes protines : 2 grdes catgories - protines qui facilitent la nutrition : 2 catgories : - protines qui aident au transport des nutriments. Protines hydrophiles qui capturent les nutriments quand ils entrent dans le priplasme et les amnent (transport : AA, sucre, vitamines) - enzyme de dgradation comme phosphatase, protase permet le recyclage des protines en fin de vie. Endonuclases : dcoupe les acides nuclique, traverse la membrane cellulaire et est utilis comme substrat. - protines qui inactivent les procds toxiques On trouve des oligosaccharides particuliers : des glycanes priplasmiques OPG dterminant de la virulence des bactries Gram - et sont des molcules signales qui permettent de percevoir les variations de lenvironnement.
Microbiologie 2 IV. Les structures facultatives. Les structures facultatives sont + ou impliques dans la pathognse. 1. Les couches de surface cristallines. = couche S Elle est forme dune seule sorte de protine et est polymrise lextrieur de la bactrie. Parfois des glycanes y sont attachs. Cette couche S est ncessaire pour la virulence. 2. La capsule. Elle est, dans la majorit des cas, un polysaccharide + ou complexe. Dans quelques rares cas, enchainement dAA. En gnrale, la synthse de la capsule est conditionnelle = synthtise dans certaines conditions environnementales. Fonction : lutte contre la dessiccation (hausse de losmolarit). Pour les pathognes, elle sert lutter contre les dfenses de lhte notamment contre la phagocytose. 3. Les flagelles. Permet de se dplacer. Facteur de virulence important. La majorit des bactries pathognes ont des flagelles. Si on leur retire ce flagelle, elles sont beaucoup moins virulentes. Structure complexe : - Gram- : - 2 disques permettent de traverser la membrane cellulaire - 2 disques permettent de traverser la membrane externe - crochet qui courbe fortement le flagelle : trs important pour le fonctionnement du flagelle procaryote - moteur aliment par le gradient lectrochimique de protons (1 000 protons / tour) - 1 filament (polymrisation dune seule protine) - Gram+ : mme chose sauf quil ny a pas de membrane externe donc 2 disque en moins. La flagelline (protine majeure du filament) est hautement antignique et permet la bactrie de se faire reprer par son hte lorsquelle est pathogne. 4. Les pilis. Ce sont des appendices dpassant lextrieur de la cellule. Les pilis sont des structures protiques beaucoup + petites en taille que le flagelle. Les pilis de type 1 et de type 4 sont des organites dattachement en surface et vont tre, en gnrale, relativement spcifiques. Pour E. coli, 200 300 pilis. Les pilis peuvent prendre naissance soit dans la membrane cellulaire soit dans la membrane externe. 6
Microbiologie 2 Les pilines Certains pilis possdent une protine polymrise, dautres plusieurs protines. Souvent lextrmit de ces pilis, il y a une protine qui est la protine de spcificit Il y a un type de pili particulier = pili sexuel joue un rle important dans la conjugaison bactrienne. Cette conjugaison est un des systmes majeur qui permet aux bactries de se transfrer lune lautre de lADN : permet le brassage de linformation gntique. Les E. coli pathognes (urinaire, mningite) ont besoins de piline pour saccrocher sur une cellule de lorgane quelle doit infecter. Ce sont des lments trs importants pour les organismes pathognes. V. Les rcepteurs des agents dltres exognes. 1. Les bactriophages. Ce sont les virus bactrie. Taille = 20 300 nm. On ne peut pas les reprer en microscopie optique (sauf exception). Mais visible en microscopie lectronique. Possde un trs grand degr de symtrie. Constitution : tte dans laquelle il y a un acide nuclique. On peut avoir une seule molcule dacide nuclique ou plusieurs. Queue (pas tjrs prsente). Dans le cas du bactriophage T4 : Tte protique renfermant lacide nuclique (ADN double brin). La tte protique est forme de sous units qui sassemblent = capsomres. Forme icosahdrique = 20 faces et 12 sommets. Possde une queue qui facilite linjection de lADN. De plus, la queue peut tre contractile. La contraction de la queue permet dinjecter le matriel gntique lintrieur de la cellule. Il y a apparemment une jonction entre la membrane cytoplasmique et la membrane externe. (Soit rapprochement des 2 membranes, soit pont protique on ne sait pas trs bien). Quelques exemples (pour E. coli): - T1, T2, T4 : possdent queue contractile 169 kb - x 174 : + petit et ne possde pas de queue. 5,4 kb - : possde une queue non contractile. Chaque espce de bactrie possde sa propre collection de bactriophage. Chaque bactriophage est spcifique dune espce. Mme certains bactriophages sont spcifiques dune seule souche pour une espce donne. Mais on dit quun phage est spcifique dune espce. La spcificit des bactriophages dpend de certains constituants de lenveloppe bactrienne. Les protines ou lipopolysaccharides ne sont pas la surface pour faciliter lentre des phages. Mais ces structures ont t dtournes de leur fonction initiale pour servir de rcepteur aux diffrents bactriophages.
Microbiologie 2 Lorsque le rcepteur du bactriophage est le lipopolysaccharide, dans ce cas, seule une partie de lespce sera infecte car il y a une variation du LPS. Dautres bactriophages reconnaissent dautres protines : - Btu B BF 23 (T4) - Fhu A T5 - Lam B Dautres structures reconnues par les bactriophages : - Pilis sexuels M13 - Flagelles : (chi) Salmonella enterica, PBS1 Bacillus subtilis. On peut considrer que toute structure en contact direct avec le milieu extrieur est probablement la cible dun ou plusieurs bactriophages. On a 3 types de bactries : - sensible : contamine puis limine par le bactriophage - rsistante : ne possde pas le rcepteur du bactriophage - immune : a le rcepteur mais possde au sein de son chromosome lADN du bactriophage et empche linfection. La majorit des bactriophages sont virulents (cd quils reconnaissent la bactrie sensible, ils se dveloppent et dtruisent la bactrie). Bactriophages temprs : ils sont capables de ne pas dtruire systmatiquement la cellule bactrienne quils ont infects. Ils ont 2 possibilits : - entre en cycle lytique : se comporte comme un phage virulent - entre en cycle lysognique : va sintgrer au gnome bactrien sans causer de tord la cellule bactrienne. Le phage va tre lintrieur du chromosome bactrien et est invisible. Le bactriophage se comporte comme un lment quelconque du chromosome. Mais lorsque la cellule bactrienne est en survie, il peut sexciser du chromosome bactrien et commencer un cycle lytique. Bactrie lysogne : quand le gnome est intgr. Dans le cas de bactriophages virulents : On considre une suspension de bactriophage et une population de bactries qui leurs sont associs. 100 bactriophages + 106 bactries : la multiplicit dinfection = 10-4. On laisse ce mlange 5 ou 10 min le temps que le bactriophage trouve une bactrie infecter. On tale ce mlange sur LB. On laisse une nuit 37C. On obtient un tapis bactrien. Les bactriophages se sont rpendus et ont contamins les bactries. Toutes les bactries proches de la bactrie initialement infecte vont lyser. Donc, on obtient des trous (sans bactrie) causs par la lyse des cellules bactriennes. Environ 100 trous. On peut suivre le devenir des bactriophages lors dune infection on le fait en milieu liquide et diffrents temps. Toutes les 5 min, on prlve la culture bactrienne et on regarde ce quil se passe.
Microbiologie 2 106 bactriophages + 5.106 bactries. Toutes les 5min, on prlve : On centrifuge la culture et on mesure le nombre de bactriophages prsents dans le surnageant. Le surnageant, on le met en contact avec des bactries sensibles pendant 10min. On ltale sur boite en milieu LB, on le laisse poser une nuit. Puis on pourra compter le nombre de bactriophage, diffrents temps, dans notre milieu de bactries. Schma 4 Phases : 1 = Phase de dclin 2 = Phase dclipse 3 = Phase explosive Dans ce graphique : on mesure le nombre de bactriophage prsent dans le surnageant. Phase de dclin : Au bout de 5 min, il ny a plu de bactriophage dans le surnageant. Tous les bactriophages se sont adsorbs la surface des bactries. Du coup, sils sont fixs la bactrie, lors de la centrifugation, ils tombent avec les bactries dans le culot. Il ny a donc rien dans le surnageant. Phase dclipse : Le bactriophage a trouv une cible. Le matriel gntique est inject lintrieur de la bactrie, ce matriel gntique sexprime en dtournant toutes les fonctions de lautre, il va rpliquer son ADN, il synthtise tout le matriel ncessaire la formation de la capside. Cette phase correspond donc : tout le temps ncessaire au bactriophage pour se multiplier et former de nouveaux bactriophages. Puis dbut de libration des nouveaux bactriophages. Phase explosive : Correspond la lyse des bactries et donc la libration massive des nouveaux bactriophages nouvellement forms. Chaque bactrie infecte par un seul bactriophage va librer une centaine de bactriophages. On a donc 100 fois plus de bactriophages la fin par rapport au dbut. La boite de ptri prsente donc des trous , pourquoi la lyse ne se rpend pas partout. Pourquoi nous nobtenons pas une boite sans aucune bactrie ? Les bactries vont se dvelopper en mme temps quelles ont acquis le phage. La bactrie lyse libre les bactriophages autour delle. Les 100 bactriophages issus de cette 1 re lyse se retrouvent proximit de la cellule bactrienne. Mais autour, il y a galement dautres bactries qui vont tre infectes par les 100 bactriophages. Pendant ce temps, les cellules bactriennes non infectes se multiplient (x2). Donc les nouveaux bactriophages infectent les nouvelles bactries etc Puis les Les bactriophages ont besoin dnergie pour pouvoir se reproduire . Cette nergie nest pas prsente si la bactrie est en phase stationnaire. Donc lorsque le milieu a t consomm entirement, les bactries sont en phase stationnaire. LADN nest pas capable de se multiplier et impossibilit de former de nouveaux bactriophages. Donc la culture nvolue pas et il reste toujours des bactries dans la boite. Poly page 26 et 27.
Microbiologie 2 2. Les bactriocines. Ce sont des protines synthtises partir dun ARN messager. Ces bactriocines sont scrtes par des bactries pour tuer dautres bactries. Les bactriocines dE. coli sont nommes : colicines. Ces colicines sont fais pour tuer dautres E. coli. En mme temps que la bactrie synthtise la colicine, elle produit une protine dimmunit. La colicine et la protine forment un complexe. Scrte dans le milieu extrieur, la colicine quitte la protine dimmunit et va pouvoir alors aller attaquer une E. coli sensible. Action possible des colicines : Les colicines vont sintgrer dans la membrane cellulaire, faire un trou. Tous les gradients vont tre bouleverss. Provoque la mort cellulaire. La colicine entre dans le cytoplasme et agit comme une nuclase = coupe lADN. La colicine considre va couper un site spcifique (ARN16S), la transcription est rendue impossible. Protines relativement grosses : 50 80 KDa. Les rcepteurs la prolicine sont les mmes que ceux reconnus par les bactriophages. Chaque structure extrieure la bactrie sert donc de rcepteur la bactriocine. Une fois que la bactriocine arrive sur sa cible, elle est dnature et traverse la membrane externe sous forme dnature. Elle est prise en charge par un complexe transmembranaire de la bactrie elle-mme. Elle va ensuite arriver la membrane cytoplasmique ou lintrieur de la cellule. La bactriocine est alors renature lintrieur de la cellule et peut infecter. Paragraphe pas retenir.
10
Microbiologie 2
Chapitre 4 : Biognse de lenveloppe bactrienne.
I. Rgulation de la composition de lenveloppe. Toutes les membranes sont des surfaces fermes qui doivent tre physiquement continues pour permettre le maintient de la cellule et sa continuit. Mais doit pouvoir grandir sans perdre sa continuit par addition de matire lipidique. Tous les constituants de lenveloppe doivent croitre de manire coordonne et au bon endroit. Chacun des lments doit tre dirig vers la bonne localisation. Chaque protine est diffrente et on doit systmatiquement les envoyer la bonne localisation. Au cours du cycle cellulaire, on a la division cellulaire et donc il faut nouveau sparer les 2 cellules filles sans rompre la continuit de lenveloppe bactrienne. Il faut permettre accroissement du peptidoglycane sans rompre la continuit du peptidoglycane. De plus, au niveau des enveloppes, les variations du milieu extrieur sont perues. Il peut donc avoir des changements importants sur la membrane en fonction des variations du milieu extrieur. Ex : Variation brutale du milieu extrieur = changement de source de carbone. 1. Lexemple de lutilisation du maltose par Escherichia coli. Schma 1 Lorsque du Glc est prsent dans un milieu de culture. Une srie de rgulateur empche lutilisation des autres sources de carbone. Donc les protines ncessaires lutilisation du maltose ne sont prsentes. Ce phnomne sappelle la diauxie. La 1re croissance est due lutilisation exclusive du glucose. Phase 2 : phase stationnaire car plu de Glc, qq minutes suffisent pour reprendre ensuite la croissance. La bactrie sadapte donc trs rapidement. En qq minutes, on passe de 0% de protines maltose 7%. La bactrie est capable de passer en quelques minutes dune alimentation Glc une alimentation maltose. II. Assemblage des protines. 1. Protines membranaires et protines scrtes. Dans lenveloppe, on a des protines solubles (hydrophiles) et des protines hydrophobes. Il faut sparer les protines qui doivent sintgrer dans une membrane des cellules qui doivent rester dans le cytoplasme. 1
Microbiologie 2 On a 2 catgories de protines : - possdent un peptide signal = partie qui indique la cellule si la protine doit tre dans la membrane ou scrte. Scrter une protine = La protine passe sous forme linaire dans la membrane mise en conformation lente de la protine car mise en conformation impossible dans le cytoplasme. - ne possde pas de peptide signal et possde une mise en conformation rapide. Correspond aux protines qui restent dans le cytoplasme. Le peptide signal est diffrent pour chaque protine, par contre il possde des caractristiques communes tous les peptides signal. Il fait de 20 40 AA en position N-term de la protine. On peut systmatiquement visualiser plusieurs parties. Au tout dbut, on a une charge positive. Ensuite 16 AA hydrophobes. Souvent au dbut de la protine mature, on a des AA neutres ou chargs ngativement. La partie dAA hydrophobes sinsre dans la membrane ce qui permet de transloquer toute la protine. Schma 2. Systme SEC : a besoin du passage de plusieurs protines dans membrane ou cytoplasme. Schma 3. Leader peptidase : protase spcifique qui va cliver spcifiquement la protine entre la fin du peptide signal et le dbut de la protine. Libration du peptide signal qui va tre dgrad. Ce clivage se fait que quand la protine est bien ancre dans la membrane. Ce schma marche que pour 2 types de cellules. Il reste voir la solution pour les protines de la membrane externe et les protines scrtes. Page 46 fig 5 Les protines de la membrane externe. Schma 4 Commence pareil que schma 3. Une structure lempche de sintgrer dans la bicouche lipidique (membrane cellulaire). Ds que la protine sort dans le priplasme, elle est prise en charge par des protines de type chaperons qui vont la protger de lagrgation, des protases. Ces protines chaperons vont conduire cette protine dnature jusqu' son lieu de rsidence dfinitive : la membrane externe. A son arrive, elle se met en conformation dans la membrane externe. Ce schma ne fonctionne pas pour les lipoprotines. Lipoprotine de Braun : Poly page 53 fig 16. Une cystine en N-term o se lie les 3 acide gras. On a notre lipoprotine compltement constitue, mais celle-ci est ancre dans la membrane cellulaire. Mais normalement elle doit se retrouver dans la membrane externe. Il va donc falloir transloquer la lipoprotine jusqu' la membrane externe implique de sortir les acides gras de la membrane cellulaire. Schma 5 2
Microbiologie 2 On a 3 tapes : - La 1re protine extrais les lipides de la membrane cellulaire. Les lipides vont tre masqus par cette protine. - Transfert de la lipoprotine vers une 2 me protine qui va acheminer la lipoprotine vers la membrane externe. - Arrive la membrane externe, une 3 me protine accepte la lipoprotine et linsre lintrieur de la membrane externe. Le systme sec est suffisant pour les bactries Gram+ car il ny a quune membrane. Gram- : Cas + difficile rsoudre. Il y a des systmes de scrtion spcifiques pour elles. Il y a 6 systmes diffrents quon peut regrouper en 2 grandes catgories : * systmes Sec-dpendants : * systmes Sec-indpendants : permet le passage de la protine en une seule tape du cytoplasme vers le milieu extrieur. Les + sophistiques contiennent 15 protines. Ex : Systme Sec - indpendant. 3 protines impliques. Schma 6. a Lensemble forme un cylindre travers lequel va pouvoir passer des protines. Une fois la protine passe, cet assemblage va pouvoir se dfaire. La protine passe dans le cylindre en forme dnature, puis se met en conformation dans le milieu extrieur. Ex : Systme Sec dpendant. Schma 6 b. La protine est prise en charge seulement dans le priplasme pour tre emmene vers lextrieur. La protine est scrte dans le priplasme o elle adopte sa mise en conformation dfinitive. Le systme de scrtion doit reconnaitre la protine qui doit tre scrte des protines qui doivent rester dans le cytoplasme. Une protine unique dans la membrane externe qui doit se mettre en multimre (Douzaine de monomres) qui permet daccommoder un pore relativement gros pour laisser passer la protine dj en conformation (nest plu linaire). Dans la membrane cellulaire, tout un ensemble de protines diffrentes. Puis plusieurs protines de liaisons qui permettent de reconnaitre la protine scrter, lamener au pore form par les protines de la membrane externe pour lemmener vers lextrieur. III.Assemblage des phospholipides. Toute la synthse des phospholipides a lieu sur la face interne de la membrane cellulaire. On forme des micelles (molcules amphiphiles). Cest ce que lon obtient lorsque lon met des phospholipides dans de leau. Il faut des conditions trs particulires pour obtenir une conformation en bicouches phospholipidiques.
Microbiologie 2 On a une composition en phospholipides identiques pour la couche ext et int dans les conditions in vitro. Les nouveau phospholipides se mettent sur la face interne mais comment passent-t-ils sur la face externe ? Ils se retournent et passe de lautre cot = hypothse simple. Mais cela serait trs couteux en nergie. On fait traverser la tte polaire une bicouche lipidique. Hypothse actuelle : il y aurait des protines particulires qui font traverser la tte polaire sans avoir de contact entre partir polaire / lipidique. Schma 7. Les phospholipides no synthtiss coulisseraient le long de la protine empchant les ttes polaires dtre en contact avec la partie hydrophobe des acides gras. On imagine un autoassemblage. Equilibre de la quantit de phospholipides entre face ext/int. Composition en phospholipides diffrente selon les endroits. Donc rgulation trs fine dans la composition en phospholipides et dans lorganisation des phospholipides dans la structure membranaire. Mais on ne sait pas ce qui permet davoir des phospholipides diffrents entre face interne et face externe. IV. Synthse de la murine. Cest une molcule qui est la cible dantibiotiques extrmement efficaces. La biosynthse de la murine est peu prs identique pour Gram et Gram+. Les diffrences sont principalement dues aux peptides qui relisent les chaines de glycane. Page 51. Fig 10 : Synthse de suffisamment de murine pour permettre laccroissement de la cellule et permettre sa futur division. Ce qui ncessite de lnergie se fait du cot cytoplasmique. Page 52 fig 11. Chez E. coli : Il y a 8 tapes qui permettent parvenir la synthse dUDP. Le 5me peptide est utilis ultrieurement dans lassemblage sera pass dans le priplasme. Lnergie forme par la coupure du 5me AA va permettre la transpepdidation ( ?). Une molcule en C55 sur lequel sont accrochs les prcurseurs du peptidoglycane. Sur C55 on greffe NAG. On obtient le module de base *. On se retrouve sur la face externe de la membrane cellulaire. Llment qui vient dtre transloqu va a donn a.
Microbiologie 2 Schma 8. Llment rose est ensuite libr lorsque n+1 est suffisant. On obtient les petites briques. On va pouvoir faire sinterconnecter la chaine avec le peptidoglycane. Page 51 fig 9 mal reprsent. Les nouveaux lments synthtiss vont devoir traverser le peptidoglycane sans le rompre ! Il faut donc des protines = PBP (Penicillin Binding Protein). Les 1res PBP sont en violet. Pour insrer la partie rose dans le peptidoglycane 2 et 4 (bleu). PbP 2 et PBP 4 ont 2 activits : - transglycosylation -transpeptidation Ces 2 activits permettent la cration dun rseau = peptidoglycane (PG). Il va quand mme y avoir des coupures transitoires du peptidoglycane pour insrer les nouvelles. Le peptidoglycane = 1 molcule. En gardant cette molcule unique, on garde les proprits du peptidoglycane. Organisation du peptidoglycane en spirale autour de la membrane cellulaire. Schma 9 Gris peptidoglycane dj prsent. Bleu : Nouvelle unit de base insrer. PBP : coupe en rose. Permet dintgrer la nouvelle unit. Accroissement de la taille du peptidoglycane. Lorsque lon intgre cette unit, il y a la fois transglycosylation et transpeptidation PBP 3 : Permet deffectuer une division cellulaire correcte entre les 2 cellules filles du peptidoglycane Septation. Intrt des antibiotiques : - on sattaque une structure vitale de la bactrie (peptidoglycane). - sattaque une structure exclusivement bactrienne pas de risque davoir de problme dans les systmes de biosynthses des eucaryotes. Ex dantibiotiques page 52 : : fosfomycine, D-cycloserine : vancomycine : bacitracine, colicine M : pnicillines, Toutes les tapes de biosynthse du peptidoglycane vont tre attaques par un inhibiteur = antibiotique. -Lactamases dtruisent le cycle -lactame et inactivent lantibiotique. Ils sont localiss dans le priplasme pour les bactries Gram- l o agissent les pnicilline.
Microbiologie 2 V. Synthse du lipopolysaccharide. Le LPS est compos de 3 parties : - lipide A synthtis au sein (face interne) de la membrane cellulaire. Puis, on allonge dessus la partie polysaccharidique du noyau (toujours face cytoplasmique). Une fois que lantigne O est activ, on le transloque (grce au cycle undecaprenyl) sur face externe de la membrane cellulaire, cest ce moment quil est accroch lextrmit des sucres. Une protine permet le retournement de lAg A face externe de la membrane cellulaire essentiel pour la biosynthse du LPS.
Microbiologie 2
Chapitre 5 : La recherche de nourriture.
I. Introduction. Dans la majorit des cas, les bactries se trouvent dans des conditions de carence nutritionnelle. Il leur faut trouver rapidement la nourriture ncessaire leur survie. Elles doivent faire a rapidement car : - nourriture limitante - bactries vivent en communaut. Dans les milieux de laboratoire et lorsque les bactries pathognes se trouvent dans lhte nourriture abondante. Comment la bactrie est capable de transporter des lments nutritifs et comment est elle capable de laccumuler dans son cytoplasme ? Ex : E. Coli On trouve 100 systmes de transports diffrents. Selon le systme de transport, il y a 1 5 protines impliques. Et il y a une soixantaine de gnes impliqus dans la motilit. On arrive environ 10% du gnome qui est dvolu au transport et au tactisme (= motilit). Schma 1 1re partie = toutes les enzymes ont une quantit de substrat suprieur ce dont elles ont besoin jusquau point rose. On a un arrt brutal en qq minutes arrive en phase stationnaire. Arriv ici, si on dose le Glc rsiduel < 1M. Tandis quau dpart (avant ajout de bactries), on est 11mM. Donc tout le Glc a t consomm. Comment les bactries peuvent-elles assurer une croissance la mme vitesse du dbut la fin ? Partie a : concentration de Glc en excs donc tout fonctionne bien Partie b : concentration de Glc infrieure aux quantits normalement ncessaires pour que cela fonctionne plein rgime. On sattendrait donc la courbe bleue. La concentration dun milieu intrieur est diffrente de la concentration dun milieu extrieur. Les systmes de transport sont capables de concentrer les nutriments lintrieur de la cellule afin de toujours maintenir une quantit de nutriment maximale mme en faible quantit de substrat. Arrt brutal car concentration basse le systme de transport ne fonctionne plu. La quasi-totalit des lments nutritifs sont absorbs par des systmes qui permettent de concentr les nutriments on va contre le gradient de concentration on utilise donc de lnergie. II. Le transport. 3 grandes fonctions du transport : * Rgulation du volume cellulaire. Implique maintien du pH, molarit tablissement et maintien de la condition ionique.
Microbiologie 2 * Extraire et concentrer les nutriments. (= les molcules ncessaires la formation dnergie et ) * Maintien des gradients ioniques de part et dautre de la membrane (Na+, K+, Ca2+) 1. Les diffrents types de transport. On va principalement parler de la concentration des nutriments. On se place chez les bactries Gram-. Les nutriments doivent traverser la membrane ext, traverser le priplasme et donc le PG (ne pose pas bcp de problmes aux molcules hydrophiles). Puis travers de la membrane cellulaire (principale obstacle de molcules hydrophiles). Dans la membrane cellulaire : On retrouve tous les systmes de transport qui doivent permettre lentre des molcules ncessaires la croissance et laisser lextrieur toutes les molcules dltres. Tous les systmes de transport seront spcifiques dune molcule ou dune famille de molcules. Les bactrier ont 5 types de transports diffrents : - La diffusion simple 1. - La diffusion facilite 2 - Le transport actif primaire 3 - Le transport actif secondaire 4 - La translocation de groupe. 5 2. La simple diffusion. Elle se fait sans intervention daucune protine. Les gaz peuvent traverser librement les membranes = diffusion simple. 3. La diffusion facilite. On parle ce moment l de transport. Une protine joue le rle de transporteur. En gnral, on a mme un transporteur strospcifique. Sans le transporteur, la molcule ne peut pas traverser la membrane toute seule. Le transporteur ne peut pas transporter de molcules autres que celle pour laquelle il a t synthtis. Le substrat est transport selon le gradient de concentration. On peut assimiler ce transporteur une enzyme. Spcificit de substrat + vitesse de transport dpend de la diffrence de concentration du substrat de part et dautre de la membrane jusqu' une vitesse limite qui correspond la saturation du transporteur. Seul le glycrol est capable dentrer par diffusion facilite. Avec ces 2 systmes (simple et facilite), on ne peut pas concentrer le substrat lintrieur de la bactrie car selon gradient. 4. Le transport actif. Le transport actif primaire, secondaire et la translocation de groupe ncessitent la prsence dnergie car ce sont des transports actifs. On tablit un gradient de concentration au lieu de lexploiter. Avantage mme basse concentration de substrat on continue daccumuler dans la cellule. 2
Microbiologie 2 Transporteur particuliers = permases. 5. Le transport actif secondaire. Lnergie = les diffrents gradients ioniques tablis de part et dautre de la membrane cellulaire. Schma 2. 2 gradients : - H+ - Na+ Selon les nutriments et le transporteur, on utilisera ou le gradient de protons ou le gradient sodique. Une protine transmembranaire (souvent dimre) va pouvoir faire entrer ou du H + ou du Na+ avec le substrat. On dissipe un gradient qui est prtabli. Au moment du transport, on nutilise pas dnergie. Par contre il va falloir recrer le gradient cest ce qui consomme de lnergie. Cest pourquoi on lappelle transport actif secondaire. Lutilisation de lnergie et le transport sont 2 tapes bien diffrentes. Le gradient de H+ est reform : - en arobiose par respiration - en anarobiose par (fermentation ?) Il existe 3 types de transport actif secondaires. Page 43 fig 1 - symport : entre dans le mme sens le substrat et ions. Entre dun anion ou molcule neutre. - antiport : la molcule H+ ou Na+ entre tandis quon expulse un ion de charge diffrente - uniport : la charge conditionne le sens de transport. Cations entrent et anions sortent. Spcificit : chaque transporteur transporte une ou une famille de molcules. Symport : le port ne veut pas dire pore !!!! Les transporteurs sont des protines qui peuvent avoir au moins 2 tats : - ferm : pour empcher lentre de molcules autres - ouvert : permet le passage, lorsque cest ncessaire, du substrat considr. Le sens du transport est dtermin par le gradient et pas par la molcule de transport. Si on pouvait inverser le gradient, les molcules passeraient dans lautre sens. On peut transporter diffrents types de molcules : - AA : Gly ou Phe, Pro par symport sodium - sucre : lactose = symport H+ - ions : antiport Na+/H+ ou antiport Na+/K+ ( ce paragraphe est inutile)
Microbiologie 2 Comment montrer le transport et laccumulation dune molcule dans une cellule ? Ajout de lactose radioactif prlvement de suspension bactrienne, filtrer, rincer par lactose non radioactif. On compte la radioactivit du filtre. Si lactose est transport augmentation de la radioactivit en fonction du temps. Comment savoir par quel moyen est transport le lactose ? On utilise des dcouplants. On introduit de petites molcules qui vont forms des pores transmembranaires qui sont suffisamment grands pour laisser passer les ions mais trop petits pour laisser passer le lactose. Donc on se retrouve avec une concentration H+ et Na+ gale de chaque cot de la membrane. On fait la mme exprience de transport. On pourra conclure de lexprience que le substrat est transport par une anion. 6. Le systme de transport sensible aux chocs osmotiques. Transport actif primaire. Sensible aux chocs osmotiques. Nexiste que chez les Gram- ou gram + ? On centrifuge les bactries en phase exponentielle. On reprend les bactries dans une solution contenant : - saccharose 20% (hypertonique). - EDTA 5mM (chlateur dions bivalents, Mg2+, Ca2+). On recentrifuge les bactries et on resuspend le culot bactrien dans une solution de MgCl 2 0,5mM 0C. On mesure alors lactivit de transport des diffrents nutriments. On introduit un substrat radioactif au milieu. On observe alors que si on ajoute du lactose radioactif, il y a transport du lactose. Alors que si on essaye de transporter du maltose ou de lhistidine pas de transport. Ici, on na donc pas de transport actif secondaire. Lajout dEDTA fait que lon pige tous les ions bivalents, donc les LPS ne sont plus bien attachs. Cration de trous temporaires dans la membrane perte de la totalit du contenu plasmique (protines, ions) qui est rpendu dans le milieu extrieur. Toutes les prootines solubles sortent dans le milieu extrieur. Dans le systme de transport, on a surement une protine soluble implique dans le transport et cest pour cela que maltose ou histidine ne sont pas transports. Cette protine est appele : protine affine priplasmique. Dans ce transport, besoin de 4 protines : - protine affine priplasmique (fixe le substrat) - 2 protines membranaires (impliqu dans le transport proprement dit) - 1 protine cytoplasmique (li la membrane, elle fournit lnergie ncessaire au transport du substrat)
Microbiologie 2 Le transport du maltose chez E. coli : (Systme transposable pour dautres substrats). Le substrat traverse la membrane ext, une fois atteint le priplasme, le maltose se fixe la protine affine priplasmique = protine Mal E. Celle-ci permet de concentrer le substrat dun facteur 5 000 par la fixation du substrat sur la protine. Mal E est donc en trs grande quantit. Cette protine conduit le substrat jusquaux transporteurs : Mal F et Mal G (qui sont prsentes en grande quantit). Puis, il y a la protine Mal K qui est en relation troite avec Mal F et Mal G. Mal K est toujours li la membrane Mal F et Mal G. Est-ce que la protine affine priplasmique acclre le transport ? Si mutant sans Mal E Il ny a plu aucun transport de maltose. Donc la protine Mal E est indispensable. Et on peut en conclure que le maltose nest pas reconnu par Mal F et Mal G. Cest le complexe Mal E-ligand qui est reconnu par Mal F et Mal G. Le ligand tout seul ne se lie pas aux transporteurs (pareil pour le maltose). Il y a donc au moins 2 conformations pour Mal E : - libre - li au ligand. Mal F et Mal G = transporteurs actifs, ils concentrent le substrat. Ex : le maltose peut tre concentr 100 000 fois. Comment et quand est utilise lnergie ? Un chercheur a trouv. Il y aurait plusieurs conformations pour Mal F et Mal G qui permettent lentre de maltose. Schma 3 Changement de conformation de Mal E qui peut alors fixer le ligand (maltose). Dans cette configuration (complexe ligand Mal E), Mal E est alors capable dinteragir avec le complexe MalF/ MalG. Le transport ncessite alors un changement de conformation de Mal F et Mal G. Le ligand se fixe que Mal F et MalG et se dtache de Mal E qui reprend sa conformation initiale. On na pas un port qui se cre, on a lentre du substrat mais il ny a jamais une ouverture totale entre intrieur et extrieur. Mal K hydrolyse une molcule dATP et cette nergie va permettre Mal F et Mal G de retrouver la conformation initiale qui les rend a nouveau comptant pour accepter le transport dune nouvelle molcule de maltose par lintermdiaire de Mal E. Lnergie dpense ce moment l est stocke sous forme conformationnelle dans les transports Mal F et Mal G. (si pas compris cette phrase pas grave). Lnergie est donc utilise durant le transport donc transport actif primaire. Dans ce cas de transport, on peut transporter nimporte quel type de molcule (sucres, vitamines). 7. La translocation de groupe (PTS). Fonctionne pour Gram+ et Gram-.
Microbiologie 2 Il ne peut transporter que des sucres. Le substrat est modifi de telle sorte quil sera directement utilisable pour sa dgradation. Source dnergie = PEP (phosphonolpyruvate) est utilis pour transporter diffrents substrat. Poly page 43 fig 3. 2 enzymes indispensables : Enzyme 1 et protine HPr PEP donne son phosphate (P) donc donne du pyruvate. Le phosphate est transport de lenzyme 1 jusque lenzyme HPr. Une fois que lenzyme 2 a fix la protine, le manitol est phosphoryl en Manitol 1 P. Cas du glucose : Le phosphate est transport jusque P III qui transfert son phosphate sur le Glc. Il y a phosphorylation du Glc en Glc 6 P. Ensuite le Glc 6 P, peut entrer directement dans la glycolyse. La 1re tape est donc lactivation en sucre phosphoryl ? Ce systme de transport permet de gagner une molcule nergtique. Pourquoi le Glc a besoin dune protine en + ? La protine III est utilise pour le transport du Glc mais est aussi utilis pour empcher les autres transports (interagit par ex avec complexe Mal G et Malf). 8. Ladsorption de substrats de masse importante. On prend lexemple de Btu B. Dans la membrane externe, on trouve la protine Btu B. Schma 4. La fixation de la vitamine B12 sur la protine Btu B ne ncessite pas dnergie. Problme : il faut de lnergie pour transloquer la vitamine sur la face interne. Il va falloir transporter lnergie jusque de la membrane cellulaire. La protine Ton B. La protine interagit avec la Btu B et va changer de conformation. Cette conformation nergtique va tre transfre la protine Btu B. Ce qui permet la translocation de la vitamine B12. 2 systmes permettent de transporter les grosses molcules : - ce systme - Porines spcifiques III.Motilit et tactismes. 1. Trouver lhabitat appropri. Les bactries arrivent dans leur habitat par hasard car elles sont dpendantes de leur environnement. Par ex : elles sont transportes par leau Donc les bactries vont pouvoir se dplacer involontairement. On parle ici dun autre dplacement plus limit : qq centimtres. 6
Microbiologie 2 Il faudra percevoir les variations du milieu extrieur. Toutes les bactries ne sont pas capables de se dplacer volontairement (toutes ne possdent pas de flagelle). Pour celles possdant des flagelles, on parle de tactisme. Tactisme car quand il y a prsence de flagelle, les bactries possdent aussi tout un systme de perception qui permet daller dans le bon sens etc Des bactries sont capables de se dplacer : - grce au flagelle - sans flagelle en glissant sur le substrat - bactries pritriches Tous ces modes de dplacement sont associs un type de tactisme. Il existe 4 types de tactisme diffrents : - chimiotactisme = tactisme qui ragit des [ ] de produits chimiques - Arotactisme = ragit aux variations de [ ] de loxygne. - Phototactisme = permet aux bactries de se dplacer vers la lumire. Certaines bactries sont mme capables de distinguer certaines longueurs dondes particulires. - Magntotactisme = permet aux cellules de se dplacer le long des lignes de force du champ magntique terrestre. Ex : E. coli : pH optimum ext = 7,2 Son tactisme cest sloigner de pH troc acide ou basique. Recherche zones tempres et vite les lumires bleues intenses. Elle est capable de se rapprocher de zones o il y a des [ ] de nourriture + leves. On observe quil y a bcp de mutants. Page 47 Il y a 3 groupes de mutants au tactisme : - 1er type de mutation = mutation dans des gnes qui codent les protines qui donnent le flagelle (partie bleu et verte) - 2me Rouge - 3me : gne qui codent des protines et qui permettent la perception des variations du milieu extrieur transmis au moteur qui dit au flagelle de tourner dans un sens ou dans lautre. 2. Mcanisme de la mobilit lie au flagelle. Comment les bactries sont capables daller dans un sens particulier ? D en partie la prsence de la forme crochet du flagelle. Lorsque les flagelles tournent dans le sens des aiguilles dune montre, on observe une structure bouriffe qui permet aux bactries de tourner sur elles-mmes. Schma 5a Lorsque les flagelles tournent dans le sens inverse des aiguilles dune montre tous les flagelles se mettent dans le mme sens pour former une sorte dhlice composite. Permet la bactrie davancer dans une direction. Schma 5b
Microbiologie 2 Permet aux bactries daller + vite = 10 20m/s. (revient 60km/h pour un humain). Les bactries avancent, sarrtent, tournent sur elles-mmes, puis repartent dans une autre direction se produit rgulirement. Lhlice ne permet pas de tourner, juste aller tout droit. Test de nage : Se fait dans 3 ou 4g / L dagar. Dans la boite, on dpose un endroit particulier les bactries (105). On attend qq heures. On commence avec un diamtre d1mm environ. On va voir les bactries qui vont se dplacer. On passe de 1mm qq cm. On voit des cercles concentriques. Il ny a pas de gradient dan ce milieu, donc pourquoi se dplace-t-elle ? Les bactries elles-mmes crs le gradient. En effet, elle consomme la nourriture la o elles sont, ensuite, il ny en a plu donc elles se dplacent etc Le gradient est donc automatiquement cre par les bactries qui consomment les nutriments et qui se multiplient. Schma 6 Rsume poly page 44. On met les bactries dans un milieu homogne. On observe au microscope : Schma 7 = Mouvement dune bactrie unie. Les ronds = bactries qui culbutent dure un dixime de seconde. Le trait = bactrie qui nage dure 1 seconde (dans un milieu homogne et donc sans gradient). Milieu avec srine : Cration dun gradient. On observe un changement de comportement des bactries. Schma 8 Toujours culbute de un dixime de sec. Mais allongement du temps de nage quand on est dans le bon sens. Par contre, lorsque que lon ne nage pas dans le bon sens (pas vers nutriments), le temps de nage se raccourci. La bactrie se dirige dans une direction compltement alatoire : - si bonne direction : allonge temps de nage - si mauvaise direction : raccourci temps de nage La bactrie analyse trs souvent son environnement. 3. Le chimiotactisme : rsultat dune marche alatoire biaise. Comment les bactries peuvent-t-elles percevoir les variations de concentrations du milieu extrieur ? 2 possibilits : - La bactrie a des capteurs de part et dautre de son corps et elle compare entre 2 zones de son corps. Mais on peut penser que la distance entre lavant et larrire de la bactrie est trop petit pour sentir des variations. Pas possible. - la bactrie peut mesurer tout moment la concentration du substrat extrieure et la comparer avec la mesure prcdente de concentration de substrat Bonne possibilit.
Microbiologie 2 Exprience : On se met dans 3 conditions diffrentes. Schma 9 * Srine = 0mM. Les bactries se dplacent, nagent, culbutent. Etat quilibr donc nage = 1 sec et culbute = 1 dixime de sec. * Srine = 0,75mM (rpartie de manire homognise). La bactrie se rend compte quelle passe de 0 0,75 mM de srine donc quelle doit allonger son temps de nage. On observe un allongement de tps de nage. La bactrie mmorise bien la concentration prcdente. *Srine = 0,24 mM (tjr jhomogne). Les bactries culbutent + souvent et le temps de nage dure moins dune seconde. La bactrie simagine quelle est tout le temps dans le mauvais sens donc elle rduit son temps de nage. Au bout dun moment, dans les conditions 2 et 3, elles retourneront 1sec / 1 dixime de seconde. 4. Nature et fonction des capteurs et de la transmission du signal. Poly page 48 Tsr, Tar et Trg = Capteurs qui peroivent les variations de concentration des molcules attractives. Tsr reconnait srine. Tar reconnait asp, et le maltone fix protine affine Ces 3 protines sont capables de mesurer la concentration. Elles sont appeles des MCP = methyl Chemoreceptor Protein. Ce sont des protine qui peuvent tre mthyles. Schma 10 Une fois que le ligand est fix transmission dinfo via dautres protines : - w = protine de connexion qui relie le rcepteur une protine essentielle = A - protine A sauto-phosphoryle et va pouvoir phosphoryler une protine diffrente : Y Elle est capable de sauto-phosphoryler (grce lhydrolyse de lATP) puis de transfrer son P sur Y. - Y : tat phosphoryl ou non. Y phosphoryl va pouvoir interagir avec le flagelle on active la culbute. Si Y est non phosphoryl nage. La nage et la culbute dpendent donc de la phosphorylation de Y et donc de celle de A. A ne peut pas dphosphoryler. - Z dphosphoryle Y. Z a une activit constante. Ltat de phosphorylation de A dpend de son interaction avec w et T.
Microbiologie 2 Milieu homogne : On observe que T est dans un tat particulier qui induit le changement de conformation de w qui permet A dtre phosphoryl qui permet de phosphoryler Y un quilibre se cr entre phosphorylation et dphosphorylation 1sec / 1dixime de sec pour un milieu homogne. Milieu haute concentration de ligand : Diminution de lautophosphorylation de A et donc diminution de la phosphorylation de Y et comme Z a une activit constante, on augmente donc le taux de Y non phosphoryl et donc on augmente la nage. + il y a de Y phosphoryl + il y a de culbute + il y a de N non phosphoryl + il y a de nage Milieu trs haute concentration de ligand : Le phnomne saccentue encore. On continue inhiber la culbute et nager + longtemps. Mais la bactrie ne va pas toujours dans le mme sens car culbute alatoire. Bactrie dans le bon sens : le ligand augmente. La bactrie se roriente dans le mauvais sens. La bactrie se rend compte que la concentration de ligand diminuer et reculbute tout de suite. Comment on arrive mmoriser la concentration ? 2 protines sont impliques : - R : activit constante. Elle rajoute des mthyl (Me) sur diffrents sites du rcepteur. Le degr de mthylation de la protine donne de la mmoire au systme. - B : existe sous de formes = phosphoryl ou non phosphoryl. Elle enlve les mthyles du rcepteur. Cest A qui phosphoryle B. Laffinit avec le ligand dpend du taux de la mthylation du rcepteur. Lorsque la concentration en ligand augmente cela conduit augmenter le degr de mthylation du rcepteur. Donc le rcepteur T va avoir une affinit diffrente pour son ligand. Comme on augmente la quantit de Me, le rcepteur devient rfractaire au ligand car le rcepteur est sur-mthyl. Sil y a une concentration suprieure de ligand il est encore + mthyl. Il devient alors rfractaire la concentration infrieure. Sil y a une concentration infrieure de ligand car mauvais sens favorise la culbute et donc B fonctionne et le rcepteur devient nouveau apte percevoir la concentration infrieure car le rcepteur est moins mthyl. Plus il y a de mthyles, + il faut une concentration leve pour que le rcepteur reconnaisse le ligand.
10
Microbiologie 2
Chapitre 6 : Communication bactrienne.
I. Les capteurs de quorum. Ce sont des systmes qui se dclenchent quand la quantit de bactrie est importante. 1. Mise en vidence. A t mis en vidence il y a un peu + de 30 ans. Mise en vidence pour la 1 re fois par des chercheurs qui travaillaient sur la production de lumire du calamar. La fluorescence des ces animaux est due la prsence de bactries dans certains endroits de leur corps. Les bactries prsentes chez des invertbrs ou des poissons = Vibrio fisheri. Raction quils font : FMNH2 + RCHO + O2 FMN + R-COOH + h (494nm) Enzyme = lucifrase. 2. Communication intraspcifique. Les bactries remplissent des cavits spcialises. Dans leau de mer, il y a une concentration de Vibrio trs faible. Lorsque le calamar nait les cavits sont vides. Il va faire rentrer dans ces cavits quelques bactries de lespce Vibrio fisheri mais pas bcp car concentration faible. Une fois que les bactries sont entres dans cet organe, le calamar leur donne des conditions de vie qui va leur permettre de se dvelopper de manire trs importante. La bactrie possde 2 flagelles ltat libre. Lorsquelle rentre dans lorgane de Vibrio, elle perd ses flagelles, change un peu de mtabolisme. Le Vibrio va pouvoir se multiplier de manire considrable grce aux conditions du milieu de lorgane. Arrive une certaine concentration, Vibrio active la lucifrase et effectue la raction le calamar devient fluorescent. Le calamar peut contrler la quantit de bactrie quil possde. Il peut contracter cet organe pour expulser les bactries sil y en a trop. Il y a mme prsence de macrophage pour que le calamar ne soit pas envahit par les bactries. La fluorescence lui sert se camoufler. La journe, il vit enfouit. La nuit, il monte en surface. Si cest une nuit avec beaucoup dtoiles, en remontant la surface, il va reprsenter une tache noire proche de la surface et tre repr. Sil est fluorescent il se confond avec la lumire. Sil ny a pas bcp de lumire expulse les bactries la concentration de bactrie diminue La raction ne se produit plus plu de fluorescence. La production de lumire est conditionne par la quantit de bactries. 3. Les bactries Gram ngatif. Pour les capteurs de quorum, on un couple de protines :
Microbiologie 2 LuxI / LuxR 2 protines indispensables pour activer la lucifrase. Schma 1. 2 milieux diffrents. Dans les 2 cas, on a apparition de lumire la mme concentration de bactrie. Donc le systme nest pas dpendant du temps de gnration mais est dpendant principalement de la concentration des bactries dans le milieu. Pourquoi une concentration donne, toutes les bactries sont fluorescentes en mme tps ? Il doit y avoir une molcule dans le milieu qui permet ce phnomne. Ide : les bactries ragissent une molcule scrte dans le milieu extrieur. Exprience : Milieu conditionn = milieu dans lequel on fait pousser les bactries, on rcupre notre culture (en phase de fluorescence), on centrifuge et rcupre le surnageant. Pour tre sr quil ny a pas de bactrie on filtre. On obtient un milieu strile dpourvu de toute trace de bactrie. A ce milieu, on ajoute des lments nutritifs. Dans ce milieu, on ajoute nouveau des bactries de type Vibrio une concentration faible. Dans ces conditions, les bactries sont fluorescentes ds le dbut. Montre que dans le milieu, il y a une molcule qui permet aux bactries de fluorescer. Ce nest pas la quantit de bactrie qui dclenche la fluorescence mais + probablement la concentration de la molcule du milieu qui permet de dclencher la fluorescence. Hypothse : Les bactries synthtisent constamment cette molcule, donc + on avance dans le temps, + la concentration de la molcule augmente. Donc la quantit de molcule scrte est proportionnelle la quantit de bactrie. Arriv une certaine concentration permet de dclencher une cascade de raction. Cette molcule scrte est un auto-inducteur (car scrt par la bactrie elle-mme). Chaque espce de bactrie possde sa propre : Acyl Homosrine Lactone (AHSL). Molcule scrte = auto-inducteur. Cest un systme qui permet la reconnaissance des bactrie de la mme espce. Schma 2 Basse concentration : A faible concentration, LuxI synthtise lauto-inducteur en permanence. Cet auto-inducteur, de part sa composition, peut diffuser travers les membrane. Donc quilibre de [AHSL] de part et dautre de la membrane. Mais LuxR nest pas capable dactiver les gnes. Forte concentration : A partir dune certaine concentration, Lux R est capable de fixer AHSL et donc dactiver les gnes. Ces gnes vont produire des lucifrases qui vont permettre de raliser la raction et donc fluorescence.
Microbiologie 2 Chez Pseudomonas aeruginosa : Nombre important de gnes virulents qui ne doivent pas tre exprimer tous au mme moment cascade de rgulation minutieuse de lexpression de gne qui mne la virulence. Pseudo a trouv un systme, elle 2 capteurs de quorum diffrents : - LasI / LasR - RhlI / RhlR Les 2 systmes sont des homologues de LuxI/LuxR RhlI va produire une AHSL qui sera reconnue par RhlR (mais pas par LasR). Il ny a pas de raction croise entre les 2. Comment peut-on avoir une cascade de rgulation ? On a plusieurs gnes de rgulation. On dit quon a 2 catgories de gnes rguls qui doivent tre exprims des moments diffrents. Ceux rgule par Las doivent tre rguls + tt que ceux rguls par Rhl. Vir I = gne exprim prcocement (Rgulateur = LasR) VirII = gne exprim + tardivement (rgulateur = RhlR) Schma 3. Basse concentration : Las I synthtise ASHL (carr sur schma) en mme temps RhlI synthtise son AHSL (triangle). Les 2 molcules dAHSL sont synthtises au mme moment. Dans les gnes rguls par LasR, on a avant rhlR. De +, on a le rgulateur LasR dj prsent dans la cellule. LasR nest pas capable dactiver rhlR et vir I. Forte concentration : Las R est alors capable de reconnaitre lAHSL correspondante qui est en concentration suffisante. On peut alors activer tous les gnes rpondants LasR. Il y a transcription des gnes de 1re catgorie des gnes de virulence. Et dbut de transcription des gnes dRhlR. Donc expression diffrentiel des gnes grce lexpression diffrentielle des rgulateurs. Au bout dun certains temps, RhlR est synthtis et est en quantit suffisante pour activer les gnes de virulence de 2me catgorie. Schma 4 La partie qui varie = bleu. 4. Bactries Gram positif. Peptide synthtis dans les bactries et scrt dans le milieu extrieur et saccumule. A une concentration suffisante de peptide dclenche lexpression. Mais le peptide est incapable de traverser la membrane tout seul. Besoin dune machinerie pour scrter le peptide dans le milieu extrieur. Une fois dans le milieu extrieur, le peptide ne rentre plu dans la bactrie il faut dtecter la concentration dans le milieu extrieur et
Microbiologie 2 transmettre linformation que les peptides sont en quantit suffisantes et permettre lexpression des gnes cibles. schma 5 Le peptide est fait partir dun petit ARNm. Cet ARNm code un petit peptide denviron 40 AA. Ce peptide va tre cliv et seul la partie rouge va tre scrte dans le milieu extrieur, les autres parties vont disparaitre. On se retrouve avec un peptide de 8/10 AA transport spcifique de transport de ce peptide. Le peptide se retrouve lextrieur. Le systme est globalement le mme que pour Gram-. Le peptide saccumule au fur et mesure que la concentration cellulaire augmente. Une protine rgulatrice doit aller activer les gnes. Dans cet tat, la protine rgulatrice ne fonctionne pas. Une protine transmembranaire est un capteur. Ce capteur va percevoir la concentration des polypeptides partir du moment o elle est suffisante. A partir de cela un mcanisme se met en place : - autophosphorylation du capteur - le capteur peut alors transmettre le P au rgulateur devient actif. - Le rgulateur va alors activer lexpression des gnes cibles. Les bactries qui utilisent ce systme : - streptococcus - bacillus subtilis 5. Communication interspcifique. On a vu les auto-inducteurs de type I. On a trouv rcemment des auto-inducteurs de type II. Structure totalement diffrente. Ces systmes peuvent tre reconnus par des bactries Gram+ et Gram-. On a montr que certaines plantes (en particulier le Varech) qui sont contamines par une bactrie particulire (Serratia Gram+). Elle digre les feuilles de Varech. On sest aperu que le Varech est devenu rsistant Serratia. Le varech scrte une molcule qui est de type AHSL reconnue par Serratia. Lorsque Serratia arrive pour coloniser, elle reconnait lAHSL, va alors exprimer des gnes de virulence au mauvais moment et ne pas russir infecter le Varech limination de Serratia.
Microbiologie 2
Chapitre 7 : Pathognie et symbiose.
I. Introduction. On se place du ct des bactries. Comment sont-elles capables de raliser une infection ? II. Les tapes de linfection. Maintenir un rservoir. La bactrie doit se trouver un endroit pour vivre ou pour survivre entre 2 infections. Cela signifie sa capacit de nuire un hte et dterminer sa capacit vivre sans son hte. Transport vers lhte. Adhrer et coloniser lhte. Multiplication dans lhte. Echapper aux dfenses de lhte. Capacit nuire son hte. Quitter son hte et retourner ltape 1.
Ces tapes sont pour les pathognes. Pour un symbionte, on remplace capacit nuire son hte par capacit apporter qqch son hte . Maintenir un rservoir. Le rservoir peut tre le milieu environnant : eau Le rservoir peut galement tre un hte intermdiaire comme un animal ou une plante, cet hte nest pas infect. Transport sur lhte. Transport via lair, leau Transfert direct entre porteur sain et hte dfinitif. Adhrer et coloniser lhte. Etape trs importante. Lorsque la bactrie entre dans lhte en quantit trs limite. Donc elle peut tre limine trs vite. En entrant dans lhte, la bactrie essaie de ne pas se faire reprer et commence se multiplier. Elle a donc besoin de nutriment. Les adhsines permettent de se fixer aux tissus de lhte infect. La bactrie doit sadapter un nouveau milieu = lhte. Multiplication dans lhte. A partir de ce moment, la bactrie dclenche la synthse de facteurs de virulence. Elle doit se dplacer et scrte des produits qui permettent de dtruire les jonctions entre les ccellules voir mme de lyser les cellules. La bactrie se multiplie et se dveloppe. Puis la bactrie quitte son hte.
Microbiologie 2 III.Les facteurs gnraux de virulence. Ces facteurs sont nombreux et assez complexes mettre en vidence. Ils sont ncessaires pour la vie lextrieur et lintrieur de lhte. Facteur de virulence = molcule bactrienne ncessaire au bon dveloppement de la bactrie. On repre ces facteurs : Lorsque lon inactive un gne et quon met ce mutant dans lhte. On regarde les facteurs de virulence dans lhte. Un gne mut conduit labsence de facteurs de virulence. La bactrie doit reconnaitre quelle est entre dans lhte, facteurs de rgulation : - temprature + leve - osmolarit + leve - abondance de nutriments - carence en fer - dtection de substances spcifiques de lhte Tous ces facteurs sont perus par la bactrie. La bactrie produit alors des facteurs de virulence. La production des facteurs de virulence est squentielle. Il existe de nombreux rgulateurs. Chaque rgulateur va rguler plusieurs facteurs de virulence diffrents. Chaque facteur de virulence va, en gnrale, tre rgul par plusieurs facteurs de rgulation (rgulateurs). IV. Les mcanismes de rsistance aux dfenses de lhte. Toutes les conditions environnementales doivent tre runies pour pouvoir dclencher correctement la virulence. Facteurs de virulence : - Adhsines - Expression et synthse de toxines. - Synthse dlments permettant de se dfendre contre les dfenses de lhte. - Capsule : permet de se dfendre contre la phagocytose. - Molcule scrte Position des gnes de virulence : - les gnes sont prsents autour - les gnes sont prsents sur un bactriophage tempr - les bactries possdent un plasmide qui porte les gnes de virulence Si le plasmide est perdu, la bactrie peut vivre lextrieur de lhte mais nest plus virulente. - tous les gnes sont regroups au mme endroit lintrieur du chromosome. Toute une zone du chromosome regroupe la totalit des gnes de virulence = Ilot de pathognicit. Ces ilots contiennent beaucoup de bases G et C. Ilots transfrables entre bactries manire pour crer de nouvelles espces pathognes V. Pathognes et symbiotes des plantes. Bactrie spectre dhte large : ne peut pas infecter une plante saine. Il faut que la plante est une blessure pour permettre la bactrie dentrer. 2
Microbiologie 2 Par contre, les b spcifiques dun hte sont capables dentrer par elles-mmes dans la plante. 1. Agrobacterium tumefaciens : la gale du collet. Agrobacterium tumefaciens est une bactrie large spectre dhte qui attaque les plantes lgumineuses. Maladie provoque = la gale du collet. En gnral, la maladie nest pas fatale la plante. Ex : vigne, plants de tomate, carottes A ltat naturel, Agt est une bactrie de la rhizosphre (proche des racines). La bactrie nest pas gnante pour la plante. Elle se multiplie et consomme des substances rejets par la plante. Elle fait partie des -protobactrie et plus particulirement de la famille des Rhizobiaces. Elle prsente un plasmide appel Ti qui porte tous les gnes de virulence. Tant que la plante est saine, elle ne peut pas tre infecte par Agt mme si prsente en grande quantit. Si la plante a une blessure, il peut y avoir colonisation de la plante par les bactries. Il faut que les bactries dtectent la blessure de la plante. Les Agt ont un flagelle et font du chimiotactisme. Une partie du chimiotactisme est ddie la dtection de la scrtion de substances spcifiques de la plante lorsquelle est blesse. Environ 10% des bactries sont capables de se dplacer + efficacement vers une blessure. Seul 10% de la population totale dAgt possde le plasmide de virulence. Toutes les bactries vont se diriger vers la blessure mais seul les 10% possdant le plasmide peroivent les substances spcifiques scrtes par la plante quand blessure. Reconnait des concentrations de 10-7 Molaire. Composs spcifiques reconnus : Ex : actosyringone joue un rle attractant pour les bactries possdant un plasmide (et jouera un autre rle + tard). Un certains nombre de permases vont tre synthtises et inclues dans les membranes dAgt va permettre de faire entrer des molcules spcifiques uniquement utilisable par Agt. Actosyringone permet la synthse dune permase spcifique dAgt. Il va y avoir htroconjugaison entre la bactrie et une cellule vgtale. Schma 1. Uniquement lADNt est transfr dans la plante. Il se retrouve dans le noyau et ce fragment sinsre au hasard dans lun des chromosomes de la plante. Tous les gnes ports dans lADNt vont ensuite sexprimer dans lADN de la plante. Ce transfert dpend dun systme de capteurs de quorum. 1. Synthse dhormones vgtales qui permettent le dveloppement des cellules vgtales qui vont conduire au dveloppement de la gale collet. On amplifie les cellules vgtales portant lADNt dans le but de permettre la cellule vgtale de produire des mtabolites qui vont tre ensuite consomms par la bactrie. Ces mtabolites sont : - agrocinopine - opines Ces molcules vont tres libres dans le milieu extrieur. Seul les Agt possdent les systmes de transport pour dgrader ces molcules et les utiliser.
Microbiologie 2 Schma 2 Schma 3 Agrobacterium radiobacter : Elle possde un plasmide particulier = pAgK84. Elle est capable de consommer les lments agrocinopine et opine mais est incapable de les faire scrter par la plante. Elle fait entrer de lagrocine dans le cytoplasme puis synthtise un analogue toxique de ladnine. Ceci est toxique pour Agt et le dtruit. Permet Agr de prendre toute la place et de consommer les nutriments. 2. Sinorhizobium meliloti : la fixation de lazote. Elle est une -protobactrie. Elle interagit avec une plante hte et tablit une symbiose. Elle scrte de lazote (N2). Elle na pas de large spectre dhte. Elle interagit avec une seule espce de plante. N2 2 NH3 16 ATP Comment reconnait-elle la plante hte ? Il faut des molcules spcifiques de la plante et de la bactrie. La plante produit tout le temps des molcules particulires mme sans tre blesse. Chaque plante produit ses propres molcules scrtes. Elle produit des molcules phnoliques. Les bactries vont percevoir ces drivs phnoliques. Et si, aux alentours de la plante, il y a la bonne bactrie les molcules de la plante vont tre reconnues par les bactries. Au moment o la bactrie peroit les molcules phnoliques scrtes par la bonne plante, elle commence alors scrter des facteurs de nodulation. Ces facteurs sont des molcules particulires = un sucre li un acide gras. Chaque espce de bactrie synthtisera son propre facteur de nodulation. La plante reconnait le facteur de nodulation Reconnaissance rciproque Cela augmente la spcificit du systme. La plante autorise alors la fixation des bactries sur les (radicelle ?) de la plante. Des rhicadsines spcifiques vont tre reconnues par des lysines spcifiques la surface de la plante. Chaque espce bactrienne a ses rhicadsines spcifiques. Le poil racinaire va alors se courber et les rhicadsines entrent dans les racines. Les bactries entrent lintrieur des cellules de la plante. Schma 4. Les bactries vont alors se multiplier de manire importante et remplir les cellules vgtales. Les bactries vont grossir de 7 fois et tre transformes en bactrodes. Ces bactrodes ne pourront plus jamais retourner ltat de bactrie. Les bactries vont tre nourries par les substances nutritives fournies par la plante. En change, la bactrie synthtise la nitrognase pour permettre la raction N2 2 NH3.
Microbiologie 2 Problme, cette nitrognase ne supporte pas loxygne. La plante dveloppe un rseau xylme/phlome pour transporter les nutriments. La chaine respiratoire des bactries fonctionne problme prsence dO2. La plante synthtise de la leghmoglobuline qui entoure compltement les bactries. La leghmoglobine : une partie synthtise par la bactrie et une autre synthtise par la plante. Le plasmide Sym contient tous les lments indispensables la symbiose. 10% seulement possdent ce plasmide. On place le plasmide dAgrobacterium dans rhizobium : Le rhizobium devient pathogne. Le plasmide Ti possde donc tous les facteurs de virulence. On place le plasmide Sym dans Agrobactrium. Agt devient une bactrie symbiotique. Elle va fabriquer des nodules mais elle sera en aucun cas capable de produire de lazote car le gne de la nitrognase est sur le chromosome et non sur le plasmide. Conclusion : La totalit des gnes de virulence ou de symbiose sont bien tous situs sur le plasmide. 3. Pseudomonas syringae : La rponse hypersensible. Il y a 2 groupes de gnes chez les bactries : - gne de virulence - gne davirulence Des gnes peuvent tre la fois de virulence et davirulence. Des gnes de rsistance vont se dclencher selon si lon est dans lhte ou non. Dans la plante hte, les gnes de virulence sont dclenchs. Dans la plante non hte, les gnes davirulence sont dclenchs et empchent la bactrie de se propager dans la plante. Rponse hypersensible. On dpose des bactries la surface de la plante non hte. La plante reconnat une agression. Les cellules disposes autour du dpt bactrien vont modifier leur mtabolisme. Entre de H + et Ca2+ et sortie de K+ forte hausse du pH extracellulaire. Les bactries naiment pas ce pH et commencent tres dtruites. Mais les cellules vgtales commencent galement tre dtruites. 1. Augmentation du pH. Destruction progressive des bactries et des cellules vgtales autour du dpt. 2. Les cellules vgtales scrtent des substances en mcanismes de dfense. 3. Les cellules vgtales dans un primtre un peu plus loign scrtent des radicaux oxygns ROS. Mais ces ROS dtruisent galement les cellules qui les produisent. Mais dtruit galement un peu les bactries.
Microbiologie 2 4. Etablissement dun cordon sanitaire pour empcher la prolifration des bactries. Dans la zone sanitaire toutes les cellules vgtales meurent. Et toutes les bactries meurent par apoptose. En parallle, on induit une rponse systmique (qui se propage toute la plante), des substances vont permettre cette zone de mieux rsister sil y a une autre infection. Pseudomonas syringae est une bactrie spcifique de lharicot. Elle cr des trous dans les feuilles. VI. Pathognes des animaux. 1. Yersinia pestis : la peste. Mort 7 jours aprs avoir t infect. 3 pandmies : 2 en Europe et 1 dans le monde entier. 3me pandmie : Hong Kong 1894. Il y a encore des morts aujourdhui environ 2000 /an. Epidmie = atteinte simultane dun grand nombre dindividus dun pays ou dune rgion par une maladie contagieuse. (Grippe, collera) Pandmie = une pidmie qui atteint les populations dune zone gographique trs tendue cd 1 ou plusieurs continents (peste, sida) Endmie = maladie particulire une rgion donne qui existe de faon quasi permanente. (Collera, paludisme) Lpidmie est accidentelle tandis que lendmie est chronique et permanente. Yersinia pestis : Entrobactrie. Elle possde un plasmide de virulence. Bactrie trs jeune car apparue il y a environ 20 000 ans. Elle est issue dune autre bactrie = Yersinia pseudotuberculosis. Yersinia pseudotuberculosis est une proche voisine de yersinia enterocolitica. En passant de Yersinia pseudotuberculosis Yersinia pestis. Yersinia pestis a perdu des gnes par rapport Yersinia pseudotuberculosis. Cette perte de gne conduirait une rgulation diffrente de gne, notamment de gnes de virulence. Ce qui la rend + agressive, + pathogne. Virulence extrme mais perd sa capacit survivre dans des milieux dnaturs (sur une table). Alexandre Yersin a mis en vidence le bacille de la peste. Lhte principal de Yersinia pestis nest pas lHomme mais le rat et la souris. LHomme est un hte intermdiaire. Les vecteurs et les htes intermdiaires obligatoires = les puces. Les puces prsentes sur les rats sautent sur lHomme et transmettent la maladie. Il existe 2 manires de transmettre la peste : Transmission par les rats / souris.
Microbiologie 2 Seule une trentaine despces de puce qui peut transmettre la peste. Lorsquune puce normale arrive sur un hte, elle le pique et se gorge de sang. Si cette puce saine pique un individu malade, elle ingurgite des bacilles de la peste. Ces bacilles vont se dvelopper dans le tube digestif de la peste, dans lsophage. Elle va compltement bloquer ce tube digestif en formant un biofilm. La puce va piquer lhte et essayer dabsorber du sang mais elle ne va pas y arriver. Donc elle va piquer continuellement pour essayer de manger. Elle rgurgite le sang aspir et donc dlivre des bactries dans lhte. La puce finit par mourir et lhte est infect son tour. Il suffit de 80 bactries pour provoquer la maladie. Cette peste sappelle la peste bubonique ou peste noire. Les bactries se dirigent ensuite vers les ganglions lymphatiques o elles se multiplient. Ces ganglions gonflent ce qui donne des bubons. Une fois que le ganglion est plein de bactrie, les bactries vont se dverser dans le sang septicmie qui entraine la mort de lindividu en 2 jours. La peste pulmonaire. Mort en 24 48h. Elle se transmet dHomme Homme. Les bactries se dissminent dans lair par ternuement, toux Lindividu sein inhale les bactries et se retrouvent dans les poumons. Les bactries se retrouvent directement dans le sang et il y a septicmie presque immdiatement. Mort trs rapide. Septicmie = Infection gnralise trs grave due la prolifration dans le sang de bactries pathognes. Les htes intermdiaires sont des rongeurs qui ne sont pas des rats ou des souris. La bactrie est capable de se maintenir pendants des annes en dormance. Elle se redclenche en prsence de lHomme, du rat ou de la souris. Ces rongeurs sont insensibles la bactrie mais peuvent la maintenir en vie = rservoir de la bactrie. 2. Shigella flexneri : la shigellose. Il en existe 3 espces mmes symptmes. Elle infecte le systme intestinal et provoque la diarrhe. Entrobactrie. Elle possde un plasmide de virulence de 220kb. Les 3 espces : - Shigella flexneri - Shigella sonnei - Shigella dysen Elles se dveloppent exactement de la mme manire et provoquent les mmes symptmes. Leurs gnomes sont un peu diffrents et se trouvent dans des rgions diffrentes. Mme si elle nest pas systmatiquement mortelle, cest une vraie pathogne. Lorsque lon est infect, on est forcment malade. 100 bactries suffisent pour tre malade. On pense mme que 10 bactries suffisent.
Microbiologie 2 Puisque cest une bactrie trs efficace, elle ne reste pas en vie trs longtemps dans un milieu non naturel. Elle doit donc se multiplier en grande quantit pour avoir une chance dinfecter. On la retrouve dans les eaux contamines. Le seul endroit o elles sont capables de se maintenir en vie part leur hte naturel est dans les produits laitiers. Ce sont des parasites intracellulaires. Elle traverse lsophage et arrive dans lestomac o elle doit rsister lacidit. Elle se dirige dans lintestin grle o elle commence effectuer son cycle de virulence. Son but est de pntrer lintrieur des cellules intestinales = cellules de la bordure en brosse. Problme : elle est incapable de pntrer dans ces cellules par le ct bordure en brosse. Il va falloir trouver une mthode alternative pour pouvoir entrer dans ces cellules. Elle entre par les cellules M de la plaque de Mayer qui ne possdent pas de bordure en brosse. Elle entre dans les cellules bordure en brosse par les faces latrales ou par en dessous. Les cellules M ne sont pas des macrophages et ne pratiquent pas la phagocytose. Shigelle a besoin de se faire phagocyter par la cellule rceptrice. Shigelle, elle-mme, dveloppe un systme pour obliger les cellules la phagocyter. Shigelle injecte sa seringue dans la cellule M. Des cellules de la bactrie entrent dans la cellule M et vont recruter lactine. Une vsicule dendocytose entre dans la cellule M grce la polymrisation de lactine et Shigelle peut entrer. Il y a dpolymrisation de lactine. Elle se multiplie alors dans la cellule M. Shigelle ressort de lautre ct de la cellule (cot oppos la lumire intestinale). Elle va se faire phagocyter par les macrophages. Une fois entre dans le macrophage, elle va induire lapoptose des macrophages. Lentre dans les macrophages provoque une haute rponse inflammatoire qui a pour consquence que de nombreux nouveaux macrophages arrivent sur le lieu de linfection ce qui augmente linflammation fragilise la membrane des cellules bordure en brosse. Les cellules bordure en brosse ne sont plus jointives car il y a le passage de cellules immunitaires. Ceci permet lentre de Shigella dans les cellules bordure en brosse. Shigella se multiplie alors dans le cytoplasme des cellules bordure en brosse. Elle est capable dliminer les bordures en brosse des cellules. Poly page 58 Shigella passe ensuite directement dune cellule bordure en brosse une autre. Elle recrute nouveau lactine et synthtise des polymres permettant de fixer lactine. Ceci lui permet de se dplacer + facilement. Diarrhe. 3. Bacillus anthracis : le charbon. = lanthrax (nom amricain). 2 formes : - spore ne se multiplie pas mais est trs rsistante. - bactrie germe = bactrie vgtative La forme vgtative a une dure de vie hors de son hte de quelques minutes quelques heures. Cette forme vgtative na aucun pouvoir pathogne.
Microbiologie 2 La forme virulente est la spore. La maladie ne peut tre dclenche qu partir de la spore. La spore est inhale. Elle germe, se multiplie et sort de lhte pour sporuler nouveau. Il y a 3 formes diffrentes de la maladie : - forme cutane : par piqure ou blessure contamination. La plus rpandue. Facile diagnostiquer et soigner. - forme intestinale : - forme pulmonaire : Pour les 2 dernires se dveloppe sans signe extrieur au dpart. Quand les 1 ers signes de la maladie apparaissent, il est trop tard car la septicmie a dj commence mort par septicmie. Cest un pathogne extracellulaire. Transitoirement, la bactrie doit passer par le macrophage. La bactrie est phagocyte son entre. Fusion avec le lysosome phagolysosome. La spore devrait disparatre car signe de germination de la spore. Elle ne germe quen milieu acide (lysosome : milieu acide). Elle se multiplie dans le phagolysosome Les cellules vgtatives se multiplient et continuent synthtiser des toxines de 2 types : - ltales : tue les cellules de lhte - facteur ddmes Elle se multiplie dans le sang septicmie, lindividu meurt. Les bactries vgtatives vont se retrouver lextrieur de lhte. Une fois hors de lhte, la sporulation des bactries vgtatives se produit. Loxygne est le signal qui indique la bactrie de sporuler. La pression partielle en oxygne est beaucoup + basse lintrieur de lhte qu lair libre. Mcanisme de dfense de cette bactrie : - Une capsule lempche dtre nouveau phagocyte par les macrophages. Polymre dacide D-glutamique. Elle nest pas antignique. - Couche S qui participe la pleine virulence de cet organisme. Il y a 2 toxines : - facteur ltal : LF - facteur ddmes : EF Ils sont inefficaces eux seul, besoin dune 3me substance : antigne protecteur (= PA). Ces 2 toxines doivent tre associes PA pour fonctionner. LF + PA EF + PA Ces 2 facteurs doivent entrer lintrieur de la cellule. Cette entre est permise par PA. Une protine de la membrane est reconnue par PA donc il y a fixation la membrane. Une protase de la cellule eucaryote va cliver en 2 parties ingales PA permet la partie la + grande de sinsrer lintrieur de la membrane de la cellule eucaryote = forme un pore. Les facteurs EF et LF peuvent alors traverser la membrane et entrer dans la cellule. Bacillus anthracis possde 2 gros plasmides :
Microbiologie 2 - pOX1 : porte les gnes de toxine, les gnes de la germination et quelques gnes du mtabolisme gnral (143 gnes) - pOX2 : porte les gnes de la capsule (85 gnes) peut tre limin. Le plasmide pOX1 est donc ici indispensable et non accessoire comme un plasmide devrait ltre.
10
Vous aimerez peut-être aussi
- Cours N°13Document6 pagesCours N°13Aicha HeniaPas encore d'évaluation
- Génétique: Les Grands Articles d'UniversalisD'EverandGénétique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Croissance BactérienneDocument15 pagesCroissance BactérienneHeraclePas encore d'évaluation
- Le hasard au cœur de la cellule: Probabilités, déterminisme, génétiqueD'EverandLe hasard au cœur de la cellule: Probabilités, déterminisme, génétiquePas encore d'évaluation
- TD 6 Les Méthodes de Mesure de La Croissance BactérienneDocument26 pagesTD 6 Les Méthodes de Mesure de La Croissance BactérienneBenyoucef AmelPas encore d'évaluation
- La Croissance BactérienneDocument16 pagesLa Croissance BactérienneL kPas encore d'évaluation
- 3cours 1 L2 SA Croissance BactérienneDocument16 pages3cours 1 L2 SA Croissance BactérienneMohamed HamelPas encore d'évaluation
- Microbio s3 TD 1Document12 pagesMicrobio s3 TD 1barguigmohamed7Pas encore d'évaluation
- Chapitre 4Document9 pagesChapitre 4khaldi aminaPas encore d'évaluation
- Mesure de La Croissance MicrobienneDocument4 pagesMesure de La Croissance Microbiennea.KPas encore d'évaluation
- BMC 10 Fiche FormationDocument10 pagesBMC 10 Fiche FormationBodering AlainPas encore d'évaluation
- Génétique Bactérienne Les Échanges Génétiques, Évolution, Plasticité Des Génomes Bactériens PDFDocument13 pagesGénétique Bactérienne Les Échanges Génétiques, Évolution, Plasticité Des Génomes Bactériens PDFkerkour-abd1523Pas encore d'évaluation
- MicrobiologieDocument5 pagesMicrobiologieInes FirasPas encore d'évaluation
- MICRORGANISMESDocument5 pagesMICRORGANISMESmatida.pinosPas encore d'évaluation
- Fiche de TDDocument3 pagesFiche de TDBodering Alain100% (1)
- Chapitre 1 Et 2Document27 pagesChapitre 1 Et 2vernetemma02Pas encore d'évaluation
- Eden + YaelDocument19 pagesEden + YaelChloé PetrolitoPas encore d'évaluation
- MCB1980 2017 Cours 3Document47 pagesMCB1980 2017 Cours 3David Frizon-PerešaPas encore d'évaluation
- Microbiologie Parasitologie Synthèse 230517 121642Document37 pagesMicrobiologie Parasitologie Synthèse 230517 121642Emeline LousbergPas encore d'évaluation
- Microbiologie F - ConvertiDocument18 pagesMicrobiologie F - ConvertiBOUMECHHOUR FatimaPas encore d'évaluation
- Chapitre 4 Culture in Vitro PDFDocument26 pagesChapitre 4 Culture in Vitro PDFyounes chouaiPas encore d'évaluation
- 7 Cours Croissance BactérienneDocument48 pages7 Cours Croissance BactérienneAnaïs Bl100% (2)
- Travail Personnel de LDocument5 pagesTravail Personnel de LekoaPas encore d'évaluation
- MICROBIOChapitre 03-04Document16 pagesMICROBIOChapitre 03-04levoa eteme fabricePas encore d'évaluation
- La Croissance BactérienneDocument30 pagesLa Croissance Bactérienneamira BENTOURA50% (2)
- Microbiologie Vétérinaire - Combinaison (s5+ s7+s7+ s8) - DZVET360-Cours-veterinairesDocument1 061 pagesMicrobiologie Vétérinaire - Combinaison (s5+ s7+s7+ s8) - DZVET360-Cours-veterinairesDZVET 360 ديزاد فات100% (2)
- S5 - Microbiologie Générale-DZVET360-Cours-veterinairesDocument197 pagesS5 - Microbiologie Générale-DZVET360-Cours-veterinairesDZVET 360 ديزاد فات100% (3)
- TD DenombrementsDocument6 pagesTD DenombrementssoufianeeslaouyPas encore d'évaluation
- Protocole Microbiologie finalDocument7 pagesProtocole Microbiologie finalEmilien DumasPas encore d'évaluation
- Chap IIDocument19 pagesChap IIDavidsonPas encore d'évaluation
- TD l2 Corrige RevisionsDocument2 pagesTD l2 Corrige Revisionspatreza100% (1)
- Chap 1 La Culture Cellulaire AnimaleDocument11 pagesChap 1 La Culture Cellulaire AnimaleLOLOPas encore d'évaluation
- Chapitre 4croissance BactérienneDocument12 pagesChapitre 4croissance BactérienneabirPas encore d'évaluation
- Micro BioDocument50 pagesMicro BioSmahane BoukhiraPas encore d'évaluation
- Introduction A La Bio CellDocument15 pagesIntroduction A La Bio CellLilou GrambertPas encore d'évaluation
- Chapitre 3 Culture Cellulaire - 231020 - 164638Document5 pagesChapitre 3 Culture Cellulaire - 231020 - 164638maria.toumi2109Pas encore d'évaluation
- Morphologie Des BacteriesDocument26 pagesMorphologie Des Bacteriesalmnaouar100% (1)
- FA La CelluleDocument7 pagesFA La Cellulenourabakayokoo10Pas encore d'évaluation
- Syllabus TP Bio Cell - 2016-2017 PDFDocument21 pagesSyllabus TP Bio Cell - 2016-2017 PDFnehemie nwayPas encore d'évaluation
- Eucaryotes Et Virologie (Version 1)Document46 pagesEucaryotes Et Virologie (Version 1)coursenligne.lille188% (17)
- TD 4 - Estimation de La Biomasse Bactérienne-1Document3 pagesTD 4 - Estimation de La Biomasse Bactérienne-1asma.guergahPas encore d'évaluation
- TD de MicrobiologieDocument8 pagesTD de MicrobiologieAmel YahiaPas encore d'évaluation
- Biologie Thomas - Xu Comte-RenduDocument4 pagesBiologie Thomas - Xu Comte-RenduThomas XuPas encore d'évaluation
- Microbiologie Générale VEDocument86 pagesMicrobiologie Générale VEkaoutar laaribiPas encore d'évaluation
- TD 2: Etude de La Sensibilité Aux Antibiotiques: Année Universitaire: 2021/2022Document5 pagesTD 2: Etude de La Sensibilité Aux Antibiotiques: Année Universitaire: 2021/2022Asmae Enneffah100% (1)
- Examen Cytobacteriologique Des Urines 2Document20 pagesExamen Cytobacteriologique Des Urines 2Anonymous 6Re7LzPas encore d'évaluation
- Cours - MAIRI Assia - Travaux Dirigés de Microbiologie Générale1Document59 pagesCours - MAIRI Assia - Travaux Dirigés de Microbiologie Générale1Benyoucef AmelPas encore d'évaluation
- Partie 1 Zoom Rapide Sur Les Microbes 2Document29 pagesPartie 1 Zoom Rapide Sur Les Microbes 2NT “NitixProd” PRODPas encore d'évaluation
- Les Systèmes de Nomenclature Et La Classification Des Groupes MicrobiensDocument7 pagesLes Systèmes de Nomenclature Et La Classification Des Groupes MicrobiensekoaPas encore d'évaluation
- Bactériologie (Version 1)Document32 pagesBactériologie (Version 1)coursenligne.lille183% (6)
- TP MM TesDocument32 pagesTP MM TesAtayi100% (1)
- CHAP 4 - Croissance BactérienneDocument26 pagesCHAP 4 - Croissance BactérienneHakima BadjPas encore d'évaluation
- TP Biologie Des MycètesDocument8 pagesTP Biologie Des Mycèteswissam khelifaPas encore d'évaluation
- Microbiologie Générale DR TOUAITIADocument35 pagesMicrobiologie Générale DR TOUAITIAعالمي الجميلPas encore d'évaluation
- Les Bactéries - NotesDocument8 pagesLes Bactéries - NoteszyteksPas encore d'évaluation
- 1-Organisation Geì - Neì - Rale de La CelluleDocument84 pages1-Organisation Geì - Neì - Rale de La CelluleMicasa- samaPas encore d'évaluation
- SPP PetchDocument12 pagesSPP PetchSofia FlkPas encore d'évaluation
- CHAPITRE 4Document11 pagesCHAPITRE 4evaaaaail124Pas encore d'évaluation
- Trafic Intracellulaire Et Compartiment CellulaireDocument24 pagesTrafic Intracellulaire Et Compartiment Cellulairecoursenligne.lille186% (7)
- ExocytoseDocument18 pagesExocytosecoursenligne.lille1100% (1)
- Mécanique Des FluidesDocument42 pagesMécanique Des Fluidescoursenligne.lille1100% (14)
- TD1 Croissance BactérienneDocument9 pagesTD1 Croissance Bactériennecoursenligne.lille1100% (9)
- Eucaryotes Et Virologie (Version 1)Document46 pagesEucaryotes Et Virologie (Version 1)coursenligne.lille188% (17)
- Hérédité Cytoplasmique PDFDocument22 pagesHérédité Cytoplasmique PDFcoursenligne.lille1100% (4)
- Bactériologie (Version 1)Document32 pagesBactériologie (Version 1)coursenligne.lille183% (6)
- Eucaryotes Et Virologie (Version 2)Document24 pagesEucaryotes Et Virologie (Version 2)coursenligne.lille1100% (3)
- Bactériologie (Version 2)Document70 pagesBactériologie (Version 2)coursenligne.lille1100% (4)
- Optique GéométriqueDocument51 pagesOptique Géométriquecoursenligne.lille1100% (1)
- TD Le TransportDocument6 pagesTD Le Transportcoursenligne.lille1Pas encore d'évaluation
- TD Mutants ThermosensiblesDocument5 pagesTD Mutants Thermosensiblescoursenligne.lille1100% (1)
- Les ChampignonsDocument8 pagesLes Champignonscoursenligne.lille1100% (6)
- AlgologieDocument17 pagesAlgologiecoursenligne.lille194% (17)
- Les ProtidesDocument17 pagesLes Protidescoursenligne.lille1100% (1)
- Fécondation Et GamétogènèseDocument17 pagesFécondation Et Gamétogènèsecoursenligne.lille1Pas encore d'évaluation
- Les GlucidesDocument21 pagesLes Glucidescoursenligne.lille1100% (3)
- Fécondation Et GamétogènèseDocument17 pagesFécondation Et Gamétogènèsecoursenligne.lille1Pas encore d'évaluation
- Transgenèse Végétale Et ApplicationsDocument16 pagesTransgenèse Végétale Et Applicationscoursenligne.lille1100% (13)
- TD Les Applications de La Fusion de ProtoplasteDocument12 pagesTD Les Applications de La Fusion de Protoplastecoursenligne.lille1100% (6)
- Le Système Nerveux VégétatifDocument15 pagesLe Système Nerveux Végétatifcoursenligne.lille1Pas encore d'évaluation
- TD Milieu de CultureDocument8 pagesTD Milieu de Culturecoursenligne.lille180% (10)
- TD Développement Des Ailes de DrosophileDocument3 pagesTD Développement Des Ailes de Drosophilecoursenligne.lille1100% (1)
- Biotechnologies VégétalesDocument27 pagesBiotechnologies Végétalescoursenligne.lille190% (30)
- L'excrétion RénaleDocument27 pagesL'excrétion Rénalecoursenligne.lille1100% (1)
- Métamorphose Des AmphibiensDocument13 pagesMétamorphose Des Amphibienscoursenligne.lille1100% (1)
- Physiologie RespiratoireDocument35 pagesPhysiologie Respiratoirecoursenligne.lille1100% (16)
- TD DiapauseDocument9 pagesTD Diapausecoursenligne.lille1100% (2)
- Mue Et Métamorphose Des InsectesDocument16 pagesMue Et Métamorphose Des Insectescoursenligne.lille1100% (7)
- 1-Structure Et Archirecture de La Membrane Plasmique NeuronaleDocument8 pages1-Structure Et Archirecture de La Membrane Plasmique NeuronalebayaPas encore d'évaluation
- Bi102 LipidesDocument112 pagesBi102 LipidesMLAN HesnaPas encore d'évaluation
- BiomembraneDocument10 pagesBiomembraneSido SidosidoPas encore d'évaluation
- 2022-2023 FC Membrane Cellulaire VFDocument38 pages2022-2023 FC Membrane Cellulaire VFmerdalorPas encore d'évaluation
- Chapitre 3 Membrane Plasmique Aspect Utrastructural 2020 2021 EnvoyeDocument79 pagesChapitre 3 Membrane Plasmique Aspect Utrastructural 2020 2021 Envoyeikram aouPas encore d'évaluation
- Taalim Pro - La Biologie Cellulaire - 2023 - CompressedDocument68 pagesTaalim Pro - La Biologie Cellulaire - 2023 - Compressedhoucine hamdonPas encore d'évaluation
- QCM CytologieDocument168 pagesQCM CytologieBenzaoui DjemanaPas encore d'évaluation
- La MembraneDocument23 pagesLa MembraneFanny GrouxPas encore d'évaluation
- Je Découvre Le Corps Humain Pour Les Nuls - Patrick GEPNERDocument320 pagesJe Découvre Le Corps Humain Pour Les Nuls - Patrick GEPNERCabinet Meddeber & Meddeber100% (5)
- Cours 1Document13 pagesCours 1Cristina Cevallos CampañaPas encore d'évaluation
- Biologie CellulaireDocument24 pagesBiologie CellulairealmnaouarPas encore d'évaluation
- Snv-Biologie Cellulaire-Les Membranes CellulairesDocument5 pagesSnv-Biologie Cellulaire-Les Membranes Cellulairesamira messoudPas encore d'évaluation
- 04 PhysiologieaDocument21 pages04 PhysiologieaFranck GilletPas encore d'évaluation
- PermeabiliteDocument35 pagesPermeabiliteSekaina DriouachPas encore d'évaluation
- LDP Es 1re c03Document5 pagesLDP Es 1re c03RAZA LUCPas encore d'évaluation
- Histologie GeneraleDocument203 pagesHistologie GeneraleAnou Kalenga100% (1)
- Cours 5-6 NotesDocument26 pagesCours 5-6 NotesCassandra MaaloufPas encore d'évaluation
- 1-La Membrane Plasmique-Structure 2022Document12 pages1-La Membrane Plasmique-Structure 2022أشرف زين الدين حسنPas encore d'évaluation
- Biochimie Structurale Des Lipides PDFDocument37 pagesBiochimie Structurale Des Lipides PDFDr.Zakaria MFTPas encore d'évaluation
- Biologie CellulaireDocument38 pagesBiologie CellulaireNa Na100% (4)
- 05 E3 Relation Structure PropriétésDocument17 pages05 E3 Relation Structure Propriétésberthou100% (1)
- Chapitre 1 - H2ODocument33 pagesChapitre 1 - H2OKarima AarabePas encore d'évaluation