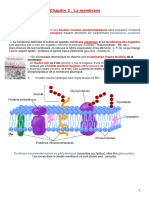
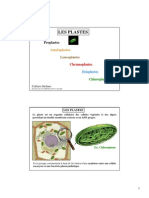
Vous aimerez peut-être aussi
- La Membrane Plasmique VF CH2Document71 pagesLa Membrane Plasmique VF CH2paul destinPas encore d'évaluation
- Résumé GCDocument4 pagesRésumé GCLilia TlbPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane PlasmiqueHou TrtyPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane Plasmiqueayman HammoudaPas encore d'évaluation
- Chapitre 2 BiomembraneDocument5 pagesChapitre 2 BiomembraneFatïma ZohraPas encore d'évaluation
- Cours 3. Membrane PlasmiqueDocument14 pagesCours 3. Membrane PlasmiqueNã BîlPas encore d'évaluation
- Cours Mod Bioch Cell Et FonctDocument73 pagesCours Mod Bioch Cell Et FonctLoubna Micha100% (1)
- Chapitre 02 La Membrane CellulaireDocument13 pagesChapitre 02 La Membrane Cellulairecqkj7sn9kxPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- 5 Membrane ErythrocytaireDocument26 pages5 Membrane ErythrocytaireBel KisPas encore d'évaluation
- La Membrane PlasmiqueDocument3 pagesLa Membrane PlasmiqueRommel EstradaPas encore d'évaluation
- Membrane Plasmique Partie 2Document45 pagesMembrane Plasmique Partie 2Oussama PanoramaPas encore d'évaluation
- Membrane PlasmiqueDocument8 pagesMembrane PlasmiqueGaston CarinePas encore d'évaluation
- Membrane PlasmiqueDocument94 pagesMembrane Plasmiquemessitaguedit10Pas encore d'évaluation
- CHAPITRE. 2 Biomembranes Cour1Document10 pagesCHAPITRE. 2 Biomembranes Cour1Joie De VivrePas encore d'évaluation
- Membrane Plasmique CH2 2022-2023Document23 pagesMembrane Plasmique CH2 2022-2023Mbaye AbouPas encore d'évaluation
- Généralité Sur La Membrane PlasmiqueDocument48 pagesGénéralité Sur La Membrane PlasmiqueOussama IdrissPas encore d'évaluation
- 02 - Revetement CellulaireDocument20 pages02 - Revetement CellulaireIssouf BerthePas encore d'évaluation
- Snv-Biologie Cellulaire-Les Membranes CellulairesDocument5 pagesSnv-Biologie Cellulaire-Les Membranes Cellulairesamira messoudPas encore d'évaluation
- Structure Des Membranes Lipidiques 1Document51 pagesStructure Des Membranes Lipidiques 1Abdellah EL AnsariPas encore d'évaluation
- 1-Structure Et Archirecture de La Membrane Plasmique NeuronaleDocument8 pages1-Structure Et Archirecture de La Membrane Plasmique NeuronalebayaPas encore d'évaluation
- Httpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFDocument12 pagesHttpsstaff - Univ-Batna2.dzsitesdefaultfilesabdessemed Samirafileschapitre 11 PDFariel.2mariemPas encore d'évaluation
- 2 Revetement Cellulaire - PPT (Mode de Compatibilité)Document21 pages2 Revetement Cellulaire - PPT (Mode de Compatibilité)Bourama KoumarePas encore d'évaluation
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- La Membrane PlasmiqueDocument51 pagesLa Membrane PlasmiquembenguemalayePas encore d'évaluation
- Biologie CellulaireDocument26 pagesBiologie CellulaireBourobou bourobou Chadrack gaelPas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- TD CorrigésDocument19 pagesTD CorrigésPierre Claver Dabou100% (1)
- Cours 3 - La Membrane PlasmiqueDocument20 pagesCours 3 - La Membrane Plasmiquemanuelakouadio405Pas encore d'évaluation
- Cours de Biologie Cellulaire L3 BMCDocument16 pagesCours de Biologie Cellulaire L3 BMCIlyasse AitPas encore d'évaluation
- CHAPITRE 3. La Membrane PlasmiqueDocument24 pagesCHAPITRE 3. La Membrane PlasmiqueRim ElmoutaoukkilPas encore d'évaluation
- Chapitre I Membrane PlasmiqueDocument17 pagesChapitre I Membrane Plasmiquesbbmxb4csdPas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Chap 3 Les Protéines Impliquées Dans La MECDocument7 pagesChap 3 Les Protéines Impliquées Dans La MECLOLOPas encore d'évaluation
- La MembraneDocument23 pagesLa MembraneFanny GrouxPas encore d'évaluation
- La Membrane Plasmique: I. GénéralitésDocument17 pagesLa Membrane Plasmique: I. GénéralitésAisSa KeRrøùùmPas encore d'évaluation
- Membrane Plasmique - WikipédiaDocument30 pagesMembrane Plasmique - Wikipédiagagnon.jonathan2007Pas encore d'évaluation
- 2eme Cour PhysiologieDocument8 pages2eme Cour Physiologiederdiri imènePas encore d'évaluation
- Membrane PlasmiqueDocument17 pagesMembrane PlasmiqueSami SamirPas encore d'évaluation
- Bio CellulaireDocument44 pagesBio CellulaireTouri Na100% (1)
- CM Biologie CellulaireDocument22 pagesCM Biologie Cellulaireleovelay34Pas encore d'évaluation
- Chap 2 MPDocument7 pagesChap 2 MPdramohandlhadjPas encore d'évaluation
- Dissertation ScientifiqueDocument5 pagesDissertation Scientifiqueseydina123ababacarPas encore d'évaluation
- Cours3 La Paroi CellulaireDocument4 pagesCours3 La Paroi CellulaireNibelMiaPas encore d'évaluation
- 3 Membrane PlasmiqueDocument53 pages3 Membrane PlasmiqueLina RazekPas encore d'évaluation
- Partie 2. Cours de BioCell 1ASNVDocument17 pagesPartie 2. Cours de BioCell 1ASNVايمن معطااللهPas encore d'évaluation
- CytologieDocument7 pagesCytologiedorissePas encore d'évaluation
- Physio1an31-2mb Plasmique NeuronaleDocument7 pagesPhysio1an31-2mb Plasmique Neuronalela raPas encore d'évaluation
- Chapitre 3 La Membrane Plasmique MedDocument6 pagesChapitre 3 La Membrane Plasmique MedDear tynaPas encore d'évaluation
- Membrane PlasmiqueDocument7 pagesMembrane PlasmiqueSara BoudjemaiPas encore d'évaluation
- Cours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESDocument102 pagesCours de Cytologie, Embryologie Et Genetique Medicale by MBAILASSEM MBAINAIDA GEORGESMãhąmãţ Hãbįb Mõųşşã ĄhmąţPas encore d'évaluation
- Groupe 8 Protéines MembranaireDocument4 pagesGroupe 8 Protéines MembranaireLounes MazariPas encore d'évaluation
- Chapitre 2 - Membrane Plasmique-Programme 2021 1Document9 pagesChapitre 2 - Membrane Plasmique-Programme 2021 1douniamhd096Pas encore d'évaluation
- Cytoplasme Bio Cell Sbaa 2024Document25 pagesCytoplasme Bio Cell Sbaa 2024habessongPas encore d'évaluation
- COURS 1 Membrane Plasmique TSBM IDocument22 pagesCOURS 1 Membrane Plasmique TSBM IMado JanicePas encore d'évaluation
- Taalim Pro - La Biologie Cellulaire - 2023 - CompressedDocument68 pagesTaalim Pro - La Biologie Cellulaire - 2023 - Compressedhoucine hamdonPas encore d'évaluation
- 2-Phosphate-La Membrane Plasmique (Exercices)Document19 pages2-Phosphate-La Membrane Plasmique (Exercices)Ay ManPas encore d'évaluation
- Structure Ultra Struc Sb1 2021 BDocument16 pagesStructure Ultra Struc Sb1 2021 BlivibotenjbPas encore d'évaluation
- La Memrane PlasmiqueDocument20 pagesLa Memrane PlasmiqueMalik NabazagaPas encore d'évaluation
- Le SommeilDocument10 pagesLe SommeilMichael GuillienPas encore d'évaluation
- Bases de La Connaissance AnatomiqueDocument9 pagesBases de La Connaissance AnatomiqueMichael GuillienPas encore d'évaluation
- RochatDocument5 pagesRochatMichael GuillienPas encore d'évaluation
- Vocabulaire de Psychanalyse Laplanche Et Pontalis Odt PDFDocument904 pagesVocabulaire de Psychanalyse Laplanche Et Pontalis Odt PDFFrixos Maravelias85% (27)
- YfcmowghzigdqasuryufDocument2 pagesYfcmowghzigdqasuryufMichael GuillienPas encore d'évaluation
- Tereno2022 attachementpsychosocialCONT 055 0013-1Document21 pagesTereno2022 attachementpsychosocialCONT 055 0013-1Michael GuillienPas encore d'évaluation
- Chapitre LN Prof p1 5Document5 pagesChapitre LN Prof p1 5Michael GuillienPas encore d'évaluation
- Histoire de La Pensée - D'homère À Jeanne D'arc (PDFDrive)Document575 pagesHistoire de La Pensée - D'homère À Jeanne D'arc (PDFDrive)Michael Guillien100% (1)
- Ds QCM Physique ChimieDocument5 pagesDs QCM Physique ChimieMichael GuillienPas encore d'évaluation
- Initiation à La Linguistique Franã Aise - 3e éd. - Sandrine Zufferey, Jacques MoeDocument350 pagesInitiation à La Linguistique Franã Aise - 3e éd. - Sandrine Zufferey, Jacques MoeMichael GuillienPas encore d'évaluation
- R Moucannas-Mazen - Le Mot Et Lâ Idã©e - Franã Ais-Arabe-Ophrys (2000) 2Document279 pagesR Moucannas-Mazen - Le Mot Et Lâ Idã©e - Franã Ais-Arabe-Ophrys (2000) 2ameer alrada100% (1)
- Chimie Generale Et Organique Par (WWW - Heights Book - Blogspot.com)Document154 pagesChimie Generale Et Organique Par (WWW - Heights Book - Blogspot.com)djamilio50% (2)
- Gazalé, Olivia by Je Taime À La Philo (Je Taime À La Philo)Document268 pagesGazalé, Olivia by Je Taime À La Philo (Je Taime À La Philo)Michael GuillienPas encore d'évaluation
- Evaluation Standadissees 1er Semestre SVT 1ere S2 2020-2021 SenegalDocument3 pagesEvaluation Standadissees 1er Semestre SVT 1ere S2 2020-2021 SenegalAmadou Makhtar Seck100% (3)
- 22955-263SC C2 2010 PDFDocument3 pages22955-263SC C2 2010 PDFFeryel ZbidiPas encore d'évaluation
- BIO CEL Chap 4 - 3 Système Endomembranaire 20-21-1Document20 pagesBIO CEL Chap 4 - 3 Système Endomembranaire 20-21-1fatimaarrach59Pas encore d'évaluation
- Memoire - VSF 4 - Copie - Copie PDFDocument110 pagesMemoire - VSF 4 - Copie - Copie PDFaliPas encore d'évaluation
- Policocopié Physiologie Végétale VraiDocument59 pagesPolicocopié Physiologie Végétale Vraibrahim chalhoubPas encore d'évaluation
- Chapitre 2 Les Constituants Chimiques de La Cellule PDFDocument10 pagesChapitre 2 Les Constituants Chimiques de La Cellule PDFAziiz BelhajPas encore d'évaluation
- La PervaporationDocument7 pagesLa Pervaporationguiguimol100% (1)
- La Membrane Biologique 1 PDFDocument11 pagesLa Membrane Biologique 1 PDFsouberePas encore d'évaluation
- CM - ToxicologieDocument40 pagesCM - ToxicologiePaul Caro0% (1)
- 6 Les Plastes CaffarriDocument14 pages6 Les Plastes Caffarriرمضان كريمPas encore d'évaluation
- Principes de Base Des Technologies A MembranesDocument39 pagesPrincipes de Base Des Technologies A MembranesLyPas encore d'évaluation
- TD ProteinesDocument30 pagesTD ProteinesSoufiane MboutayebPas encore d'évaluation
- Ex Biol 12Document13 pagesEx Biol 12Abdillahi MahadPas encore d'évaluation
- L'absorption Des Médicaments: DR Bekhti Année 2020-2021Document19 pagesL'absorption Des Médicaments: DR Bekhti Année 2020-2021safia bent aliPas encore d'évaluation
- Examen BC Session Normale 2021Document4 pagesExamen BC Session Normale 2021xPas encore d'évaluation
- Physio VégétaleDocument92 pagesPhysio VégétaleBrahimPas encore d'évaluation
- QCM Membrane Plasmique 2022Document12 pagesQCM Membrane Plasmique 2022franck brice bado100% (1)
- CHM-1007 11511Document8 pagesCHM-1007 11511tabla wiePas encore d'évaluation
- L1 BPC Part 1 Cel Excitables VVW v2-8 Light CM1Document29 pagesL1 BPC Part 1 Cel Excitables VVW v2-8 Light CM1remi.giryPas encore d'évaluation
- 8.annales IFMKDocument15 pages8.annales IFMKfy46kcjbchPas encore d'évaluation
- 16292971Document185 pages16292971Mohamed Wassim M'henniPas encore d'évaluation
- UE31 BPH03 Boi Memb 2Document65 pagesUE31 BPH03 Boi Memb 2Christian DuboisPas encore d'évaluation
- Th1 Ch2 Act3 Microscopie + Membrane PlasmiqueDocument5 pagesTh1 Ch2 Act3 Microscopie + Membrane PlasmiqueMusab BilginPas encore d'évaluation
- Biocell 1 - Cytologie (Juin 2019, Laura T)Document220 pagesBiocell 1 - Cytologie (Juin 2019, Laura T)Daniela RiosPas encore d'évaluation
- Biosynthèse Des Lipides MembranairesDocument7 pagesBiosynthèse Des Lipides Membranaireslabidiridha8Pas encore d'évaluation
- BIOLOGIE 11è SEDocument37 pagesBIOLOGIE 11è SESawoulou celest Dopavogui100% (11)
- Medecine Batna-ProgramDocument65 pagesMedecine Batna-ProgramNesrine RAPas encore d'évaluation
- Lecon 4 1S2 2024Document1 pageLecon 4 1S2 2024traoremoussatraore89Pas encore d'évaluation
- Argumentaire Spherocytose Hereditaire Et Autres Anemies Hemol Par Anomalie MB GRDocument66 pagesArgumentaire Spherocytose Hereditaire Et Autres Anemies Hemol Par Anomalie MB GREveline KoivoguiPas encore d'évaluation
- Quiz Mitochondrieetchloroplasterevision2tsbcDocument5 pagesQuiz Mitochondrieetchloroplasterevision2tsbcAldrin KiongPas encore d'évaluation