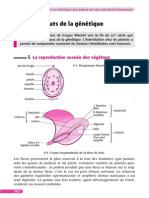
Vous aimerez peut-être aussi
- Au-delà de l'ADN: La Révolution Épigénétique: "Des Mécanismes Cellulaires aux Facteurs Environnementaux : Comment l'Épigénétique Façonne Notre Destin Biologique et les Implications pour la Santé, le Comportement et l'Avenir de la Recherche"D'EverandAu-delà de l'ADN: La Révolution Épigénétique: "Des Mécanismes Cellulaires aux Facteurs Environnementaux : Comment l'Épigénétique Façonne Notre Destin Biologique et les Implications pour la Santé, le Comportement et l'Avenir de la Recherche"Pas encore d'évaluation
- Chap2 PapierDocument26 pagesChap2 PapierNoahPas encore d'évaluation
- F - Notions GénétiquesDocument7 pagesF - Notions GénétiquesCamilaa BchPas encore d'évaluation
- 4-Nature Et Fonction Des GènesDocument16 pages4-Nature Et Fonction Des GènesWi SsalPas encore d'évaluation
- Rappels Indispensables de GénétiqueDocument10 pagesRappels Indispensables de Génétiquebakiclio5Pas encore d'évaluation
- Revsion Genetique Des DiploidesDocument4 pagesRevsion Genetique Des Diploidesle sage0% (1)
- Chapitre 2Document49 pagesChapitre 2Rahma HosniPas encore d'évaluation
- Des Débuts de La Génétique Aux Enjeux ActuelsDocument8 pagesDes Débuts de La Génétique Aux Enjeux ActuelsJosé Ahanda NguiniPas encore d'évaluation
- Geno Meet ExpressionDocument11 pagesGeno Meet ExpressionOwassiPas encore d'évaluation
- BIOLOGIE Terminales SE SS SMDocument102 pagesBIOLOGIE Terminales SE SS SMOusmane Sylla100% (2)
- Cours Complet 2022Document34 pagesCours Complet 2022Hhh GbbPas encore d'évaluation
- Inform Génétique Part 2Document63 pagesInform Génétique Part 2Ferssiwi OmarPas encore d'évaluation
- Presentation - Genetique - Formelle Pharmacie 20 21 Lois - de - MendelDocument33 pagesPresentation - Genetique - Formelle Pharmacie 20 21 Lois - de - Mendelsmart.creativity22Pas encore d'évaluation
- Les Eléments MobilesDocument34 pagesLes Eléments Mobileskerkour-abd1523100% (2)
- $R7FKSLTDocument10 pages$R7FKSLToumaimabenyoussof03Pas encore d'évaluation
- Cours 1 2021 ÉtudiantsDocument53 pagesCours 1 2021 ÉtudiantsdhiadhayoutadhayouPas encore d'évaluation
- De L - ADN À La Protéine - EducloudDocument92 pagesDe L - ADN À La Protéine - EducloudsananasPas encore d'évaluation
- Notions Amélioration PlantesDocument4 pagesNotions Amélioration PlantesPatrick EbaPas encore d'évaluation
- Cours Carte GnetiqueDocument70 pagesCours Carte GnetiquesambaPas encore d'évaluation
- Résumé M1 Protection Des Végétaux Genetique Des Populations Et Modélisation Compressed 1pdf - IoDocument19 pagesRésumé M1 Protection Des Végétaux Genetique Des Populations Et Modélisation Compressed 1pdf - IoKhalid Sidi AbdouPas encore d'évaluation
- Chapitre 2 - Med 1 - Bio MolDocument12 pagesChapitre 2 - Med 1 - Bio Molyves100% (1)
- ADNcoursDocument18 pagesADNcourslilaova12456eraPas encore d'évaluation
- 15.02.2016 La RéplicationDocument10 pages15.02.2016 La RéplicationEaPas encore d'évaluation
- Génétique SynthèseDocument55 pagesGénétique SynthèseArnaud LiebinPas encore d'évaluation
- Chap 1 - Introduction Et Lois de MendelDocument10 pagesChap 1 - Introduction Et Lois de MendelmuratPas encore d'évaluation
- Définitions SM 2020-1Document4 pagesDéfinitions SM 2020-1Hamza AmjahdiPas encore d'évaluation
- La Cartographie Génétique COURSDocument62 pagesLa Cartographie Génétique COURSAdli OthmanePas encore d'évaluation
- Cours - Unite I - Lois Statistiques - Lydex - 2020 - 2021 - Copie Prof - 2eme - SMDocument31 pagesCours - Unite I - Lois Statistiques - Lydex - 2020 - 2021 - Copie Prof - 2eme - SMhamza.aitounrar.96Pas encore d'évaluation
- ADN ARN Et Protéines 5GTSB - ShortDocument27 pagesADN ARN Et Protéines 5GTSB - Shortchelseaakame90Pas encore d'évaluation
- Blog 2Document1 pageBlog 2tozman1234gta5Pas encore d'évaluation
- Rappel Génétique 2SC PDFDocument28 pagesRappel Génétique 2SC PDFnourPas encore d'évaluation
- Biomol IFSDocument42 pagesBiomol IFSbertin adaPas encore d'évaluation
- (Suite) GENETIQUE 1BACSMDocument11 pages(Suite) GENETIQUE 1BACSMnnadanada2901Pas encore d'évaluation
- 3 Cartographie GénétiqueDocument12 pages3 Cartographie Génétiquebouchakour meryemPas encore d'évaluation
- Bilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueDocument3 pagesBilan Chapitre 2 La Replication de l' Adn Et Variabilite GenetiqueAymane JiraouiPas encore d'évaluation
- Exb Genetique RangersDocument2 pagesExb Genetique RangersM'bouillé SissokoPas encore d'évaluation
- La Génétique: Encadré Par: Présenter Par: Mme - AthmaniDocument10 pagesLa Génétique: Encadré Par: Présenter Par: Mme - AthmaniFatima ZohraPas encore d'évaluation
- Cours Bio - Mol 4emeDocument34 pagesCours Bio - Mol 4emeIBTIHEL MENZERPas encore d'évaluation
- Notion de L Information Genetique Cours 2 3Document8 pagesNotion de L Information Genetique Cours 2 3Malak MalakchakdiPas encore d'évaluation
- SVT Chapitre 1Document4 pagesSVT Chapitre 1scoccimarro.sarahPas encore d'évaluation
- Biologie Harim MalakDocument43 pagesBiologie Harim MalakMohamed SylvestrePas encore d'évaluation
- GénetiqueDocument45 pagesGénetiqueohdaeso100% (4)
- Transmission Des CaractèresDocument36 pagesTransmission Des CaractèresYaya TraoréPas encore d'évaluation
- Glossaire GénétiqueDocument13 pagesGlossaire GénétiqueBemmoussat AsmaPas encore d'évaluation
- Le Brassage Génétique Par La Méiose Et La FécondationDocument15 pagesLe Brassage Génétique Par La Méiose Et La Fécondationbouchakour meryemPas encore d'évaluation
- Cours TSE Partie1vf-1Document14 pagesCours TSE Partie1vf-1Jœël Crèspœ KhæmænœPas encore d'évaluation
- L'expression de L'information GénétiqueDocument15 pagesL'expression de L'information GénétiqueAMINE 16Pas encore d'évaluation
- La Réplication de l'ADN - Fiche de Cours - SVT - SchoolMouv5 PDFDocument8 pagesLa Réplication de l'ADN - Fiche de Cours - SVT - SchoolMouv5 PDFjuini rawiaPas encore d'évaluation
- Local Media3584551603709123337Document6 pagesLocal Media3584551603709123337Da RyaPas encore d'évaluation
- Génétique MoléculaireDocument30 pagesGénétique MoléculairemotorkinaPas encore d'évaluation
- Génétique Chap1Document17 pagesGénétique Chap1Zakaria Sadki100% (3)
- 3 Genetique Mendelienne 1Document59 pages3 Genetique Mendelienne 1charaf eddinePas encore d'évaluation
- Transformation - Copie 1Document16 pagesTransformation - Copie 1Memoire 2223Pas encore d'évaluation
- Medecine-Intro Genetique MonohybridismeDocument23 pagesMedecine-Intro Genetique MonohybridismedzsergioPas encore d'évaluation
- Polycopier Génétique Des EucaryotesDocument72 pagesPolycopier Génétique Des EucaryotessabrinaPas encore d'évaluation
- Reproduction Cellulaire 2023Document23 pagesReproduction Cellulaire 2023Jonathan Sola FuentesPas encore d'évaluation
- 11-Organistaion Du Génome HumainDocument38 pages11-Organistaion Du Génome HumainMellifluous LullabyPas encore d'évaluation
- Mes gènes, mon identité ?: Comprendre la génétique et ses enjeuxD'EverandMes gènes, mon identité ?: Comprendre la génétique et ses enjeuxPas encore d'évaluation
- Génétique: Les Grands Articles d'UniversalisD'EverandGénétique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Diapositives Fonctions Vtamines Et CoenzymesDocument12 pagesDiapositives Fonctions Vtamines Et CoenzymesAdmi BraccPas encore d'évaluation
- Synapses PDFDocument17 pagesSynapses PDFAdmi BraccPas encore d'évaluation
- Devenir Le Meilleur de Soi-Même - Abraham MaslowDocument497 pagesDevenir Le Meilleur de Soi-Même - Abraham MaslowKouakou Kouamé Rodrigue91% (11)
- Dup (01) Soc 180 (Questions - Reponses)Document8 pagesDup (01) Soc 180 (Questions - Reponses)Admi Bracc100% (1)
- La Cavitation Chez Les PlantesDocument9 pagesLa Cavitation Chez Les PlantesAdmi BraccPas encore d'évaluation
- SVT FASICULE 3eme 1Document56 pagesSVT FASICULE 3eme 1Methzo ThiamPas encore d'évaluation
- DIPLO Complementation PDFDocument4 pagesDIPLO Complementation PDFAdmi BraccPas encore d'évaluation
- Exercice 2 TD Haplophase PDFDocument1 pageExercice 2 TD Haplophase PDFAdmi BraccPas encore d'évaluation
- Pourquoi Active Windows Avce Kmspico Et Comment PDFDocument4 pagesPourquoi Active Windows Avce Kmspico Et Comment PDFAdmi BraccPas encore d'évaluation
- Cas Des Ascomycètes PDFDocument3 pagesCas Des Ascomycètes PDFAdmi BraccPas encore d'évaluation
- 2021 BOT262 CELL VEGET - CHAP3bis - Aout 09Document4 pages2021 BOT262 CELL VEGET - CHAP3bis - Aout 09Admi BraccPas encore d'évaluation
- Chp2 Partie C.TDDocument2 pagesChp2 Partie C.TDAdmi BraccPas encore d'évaluation
- Haplophase - Transmission Simultanée de Plus de Gènes PDFDocument2 pagesHaplophase - Transmission Simultanée de Plus de Gènes PDFAdmi BraccPas encore d'évaluation
- DIPLO - Dominance Et Propriétés PDFDocument4 pagesDIPLO - Dominance Et Propriétés PDFAdmi BraccPas encore d'évaluation
- Situation D'évaluation: ContexteDocument2 pagesSituation D'évaluation: ContexteAdmi BraccPas encore d'évaluation
- Caractéristiques Distinctives Entre Heredité Chromosomique Et Non Chromosomique PDFDocument2 pagesCaractéristiques Distinctives Entre Heredité Chromosomique Et Non Chromosomique PDFAdmi BraccPas encore d'évaluation
- Complementation PDFDocument5 pagesComplementation PDFAdmi BraccPas encore d'évaluation
- Absorption de L'Eau Par La PlanteDocument8 pagesAbsorption de L'Eau Par La PlanteAdmi BraccPas encore d'évaluation
- Mise en Evidence, Symptomes Et Causes Des Carences en Elements MinerauxDocument9 pagesMise en Evidence, Symptomes Et Causes Des Carences en Elements MinerauxAdmi BraccPas encore d'évaluation
- Schema General de L'Absorption & Methodes D'EtudeDocument6 pagesSchema General de L'Absorption & Methodes D'EtudeAdmi BraccPas encore d'évaluation
- Epreuve de Physique Premiere C Annee 2020 Lycee Scientifique deDocument3 pagesEpreuve de Physique Premiere C Annee 2020 Lycee Scientifique deAdmi BraccPas encore d'évaluation
- Continu It ÉDocument150 pagesContinu It ÉPriscard100% (2)
- Bot 134: Physiologie de La Nutrition Minérale Et de L'alimentation en Eau Chez Les VégétauxDocument4 pagesBot 134: Physiologie de La Nutrition Minérale Et de L'alimentation en Eau Chez Les VégétauxAdmi BraccPas encore d'évaluation
- DIPLO - Epistasie Expliquée PDFDocument6 pagesDIPLO - Epistasie Expliquée PDFAdmi BraccPas encore d'évaluation
- Carto 6Document20 pagesCarto 6Admi BraccPas encore d'évaluation
- Fiche Explicative de La Leçon - Caryotypes - NagwaDocument9 pagesFiche Explicative de La Leçon - Caryotypes - NagwaAdmi BraccPas encore d'évaluation
- Haplophase - Différence Due À Un Seul Gène 1Document5 pagesHaplophase - Différence Due À Un Seul Gène 1Admi BraccPas encore d'évaluation
- Bepc Blanc Régional 2023 Zone AboboDocument21 pagesBepc Blanc Régional 2023 Zone AboboAdmi Bracc100% (1)
- Guide en Litterature Toute SerieDocument14 pagesGuide en Litterature Toute SerieAdmi BraccPas encore d'évaluation
- TP Controle Optimal PDFDocument29 pagesTP Controle Optimal PDFHamza El-houariPas encore d'évaluation
- Examen Corrigé Marketing - Questionnaire À Choix Multiple (QCM)Document7 pagesExamen Corrigé Marketing - Questionnaire À Choix Multiple (QCM)Mar IemPas encore d'évaluation
- 1 - Cours2 Hydraulique Et Pneumatique (2éme Année Électromécanique)Document16 pages1 - Cours2 Hydraulique Et Pneumatique (2éme Année Électromécanique)Mohamed Hechmi JERIDIPas encore d'évaluation
- Corrige Serie5Document4 pagesCorrige Serie5Chef FatimaPas encore d'évaluation
- Formation - PDF - Robotique - Mécatronique PDFDocument9 pagesFormation - PDF - Robotique - Mécatronique PDFHachicha SaberPas encore d'évaluation
- 5-Focales-Episode-2-Planification Et RégulationDocument42 pages5-Focales-Episode-2-Planification Et RégulationTaffyd MaertinsPas encore d'évaluation
- Secours en Cours de RouteDocument32 pagesSecours en Cours de Routeantonio59100Pas encore d'évaluation
- Transformateurs MonophasésDocument51 pagesTransformateurs MonophasésDo OuPas encore d'évaluation
- CH1 (Introduction Au SI) PDFDocument17 pagesCH1 (Introduction Au SI) PDFEL OUAZZANI PRINTPas encore d'évaluation
- Examen Blanc Physique Chimie Bac DDocument4 pagesExamen Blanc Physique Chimie Bac Dkouassi hermann ulrich koffi100% (1)
- Programme Maths cm2Document1 pageProgramme Maths cm2gorguy100% (1)
- DRAFT - Cours OCO - Part1Document32 pagesDRAFT - Cours OCO - Part1Khennoune WissamPas encore d'évaluation
- Laboratoire Biolab Bamako - Recherche GoogleDocument1 pageLaboratoire Biolab Bamako - Recherche GoogleAlou CoulibalyPas encore d'évaluation
- Brochure Nissan Urvan N350 15 PlacesDocument14 pagesBrochure Nissan Urvan N350 15 PlaceskonePas encore d'évaluation
- World of Warcraft - Wrath of The Lich King - Arthas, My SonDocument4 pagesWorld of Warcraft - Wrath of The Lich King - Arthas, My SonAdrian Angel LagrecaPas encore d'évaluation
- Método de Violino - CajatiDocument41 pagesMétodo de Violino - CajatiWalmir PaiãoPas encore d'évaluation
- Chapitre 3 Ressources Réserves PlanificationDocument10 pagesChapitre 3 Ressources Réserves PlanificationAdlanePas encore d'évaluation
- 2M120 TD 2016 ch1 Codes CorrecteursDocument2 pages2M120 TD 2016 ch1 Codes CorrecteurskokiPas encore d'évaluation
- Biochimie 08 - Réplication Et Réparation Des Erreurs AssociéesDocument14 pagesBiochimie 08 - Réplication Et Réparation Des Erreurs AssociéesNathan CohenPas encore d'évaluation
- Serveur D'impression Sous Windows 2003Document6 pagesServeur D'impression Sous Windows 2003Issam BammouPas encore d'évaluation
- EKIP360 PrestationsTarifications LFRDocument30 pagesEKIP360 PrestationsTarifications LFRtouzanimahdiPas encore d'évaluation
- RIB CopieDocument1 pageRIB CopieAbdoulaye CisséPas encore d'évaluation
- Les Approches Managériales Basées Sur Les Parties PrenantesDocument4 pagesLes Approches Managériales Basées Sur Les Parties PrenantesNohaila BencherquiPas encore d'évaluation
- ContexteDocument2 pagesContextehibaouafi502Pas encore d'évaluation
- Exercice 3Document1 pageExercice 3test lili100% (1)
- Les Corriges Essai District Kanel Jan 2024 - 061004Document8 pagesLes Corriges Essai District Kanel Jan 2024 - 061004souleymanehousnaPas encore d'évaluation
- QP1C General 1 PDFDocument5 pagesQP1C General 1 PDFSamson KlouganPas encore d'évaluation
- Catalogue OKOFEN 2018 2019Document68 pagesCatalogue OKOFEN 2018 2019joseph38Pas encore d'évaluation
- MicrobiologieDocument5 pagesMicrobiologieInes FirasPas encore d'évaluation
- Manuel C2Document140 pagesManuel C2Daniel AndriamananaPas encore d'évaluation