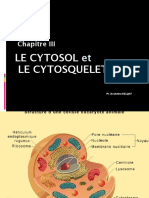
Vous aimerez peut-être aussi
- Membranes cellulaires: Les Grands Articles d'UniversalisD'EverandMembranes cellulaires: Les Grands Articles d'UniversalisPas encore d'évaluation
- Chapitre Vi Paroi PectocellulosiqueDocument9 pagesChapitre Vi Paroi Pectocellulosiqueleonce sedjroPas encore d'évaluation
- Champignons: Les Grands Articles d'UniversalisD'EverandChampignons: Les Grands Articles d'UniversalisPas encore d'évaluation
- Groupe2 Composes ParietauxDocument32 pagesGroupe2 Composes ParietauxCheick ahmedPas encore d'évaluation
- Cours 2 de Biologie Végétale: Présentés Par Mme MELLAH RDocument67 pagesCours 2 de Biologie Végétale: Présentés Par Mme MELLAH RMellah RabhaPas encore d'évaluation
- Cours 2-La Paroi PectocellulosiqueDocument66 pagesCours 2-La Paroi PectocellulosiqueMellah Rabha100% (4)
- 184 Cellulose 00Document24 pages184 Cellulose 00anwarelhilaly559Pas encore d'évaluation
- Cours3 La Paroi CellulaireDocument4 pagesCours3 La Paroi CellulaireNibelMiaPas encore d'évaluation
- Structure Et Physiologie Des Eucaryotes 2018-2019-L2S4Document7 pagesStructure Et Physiologie Des Eucaryotes 2018-2019-L2S4Original ZeyePas encore d'évaluation
- Chapitre1 Biochimie Cellulaire Et FonctionnelleDocument12 pagesChapitre1 Biochimie Cellulaire Et Fonctionnelledraboissouf174Pas encore d'évaluation
- Chapitre 2 BiomembraneDocument5 pagesChapitre 2 BiomembraneFatïma ZohraPas encore d'évaluation
- La Matrice Extracellulaire 2015-2016 PDFDocument3 pagesLa Matrice Extracellulaire 2015-2016 PDFAnonymous wVr908lBcK100% (2)
- La Paroi SquelettiqueDocument6 pagesLa Paroi SquelettiqueSara BoudjemaiPas encore d'évaluation
- 1.cours Biologie CellulaireDocument63 pages1.cours Biologie Cellulaireبدر الذهابيPas encore d'évaluation
- Résumé GCDocument3 pagesRésumé GCLilia TlbPas encore d'évaluation
- 04 PhysiologieaDocument21 pages04 PhysiologieaFranck GilletPas encore d'évaluation
- Généralité Sur La Membrane PlasmiqueDocument48 pagesGénéralité Sur La Membrane PlasmiqueOussama IdrissPas encore d'évaluation
- 1-Structure Et Archirecture de La Membrane Plasmique NeuronaleDocument8 pages1-Structure Et Archirecture de La Membrane Plasmique NeuronalebayaPas encore d'évaluation
- Cours BCG UFR2SDocument19 pagesCours BCG UFR2SSirandou DiaoPas encore d'évaluation
- Chapitre 1 Histologie-ConvertiDocument10 pagesChapitre 1 Histologie-ConvertiMond DayPas encore d'évaluation
- Présentation.41.ppt PAROIDocument12 pagesPrésentation.41.ppt PAROIRêda SlimiPas encore d'évaluation
- 2021 Bot262 Cell Vegetale - Semain3bis Aout 16Document12 pages2021 Bot262 Cell Vegetale - Semain3bis Aout 16Admi BraccPas encore d'évaluation
- ExtraitDocument8 pagesExtraitAshraf Ouro-adohiPas encore d'évaluation
- Cours Biologie Végétale Chap 1 2 3Document37 pagesCours Biologie Végétale Chap 1 2 3Nabil holmes100% (6)
- Les Matrices ExtracellulairesDocument18 pagesLes Matrices ExtracellulairesAmï NãPas encore d'évaluation
- BiocellPCEM12002 2003 Cours MaJ 2003 04 11Document58 pagesBiocellPCEM12002 2003 Cours MaJ 2003 04 11Gilbert ThierryPas encore d'évaluation
- La CelluleDocument26 pagesLa CellulesaadiaPas encore d'évaluation
- Membrane Plasmique - WikipédiaDocument30 pagesMembrane Plasmique - Wikipédiagagnon.jonathan2007Pas encore d'évaluation
- COURS BCF EtudiantsDocument22 pagesCOURS BCF Etudiantsamine bedjaPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane Plasmiqueayman HammoudaPas encore d'évaluation
- Cours Chapitre 2 Membrane PlasmiqueDocument5 pagesCours Chapitre 2 Membrane PlasmiqueHou TrtyPas encore d'évaluation
- La CelluleDocument6 pagesLa Celluleikram dahdouhPas encore d'évaluation
- (PACES) Physiologie Humaine by Hervé GuénardDocument1 371 pages(PACES) Physiologie Humaine by Hervé Guénardsam egoroffPas encore d'évaluation
- Le CytosqueletteDocument9 pagesLe CytosqueletteAmï NãPas encore d'évaluation
- Cours de Biologie Cellulaire L3 BMCDocument16 pagesCours de Biologie Cellulaire L3 BMCIlyasse AitPas encore d'évaluation
- Bacterio 1Document16 pagesBacterio 1maryPas encore d'évaluation
- Le Tissu Conjonctif Non Spécialisé WordDocument10 pagesLe Tissu Conjonctif Non Spécialisé WordAbdoulaziz SidilaminePas encore d'évaluation
- 2eme Cour PhysiologieDocument8 pages2eme Cour Physiologiederdiri imènePas encore d'évaluation
- Chapitre 2Document7 pagesChapitre 2ricky marvingPas encore d'évaluation
- Exposé Biochimie Cellulaire Et FonctionnelleDocument12 pagesExposé Biochimie Cellulaire Et FonctionnelleairaPas encore d'évaluation
- 02 Revetement-CellulaireDocument6 pages02 Revetement-CellulaireSagno AgnèsPas encore d'évaluation
- 07 - Adhérence CellulaireDocument6 pages07 - Adhérence CellulaireFairy MedPas encore d'évaluation
- Le Cytosquelette - PpsDocument14 pagesLe Cytosquelette - PpsnasredinnePas encore d'évaluation
- Membrane PlasmiqueDocument94 pagesMembrane Plasmiquemessitaguedit10Pas encore d'évaluation
- CHAPITRE I-Introduction-BVDocument8 pagesCHAPITRE I-Introduction-BVhanane boutajPas encore d'évaluation
- Solution Des Exercices PDFDocument21 pagesSolution Des Exercices PDFGabin DossouPas encore d'évaluation
- Membrane Plasmique CH2 2022-2023Document23 pagesMembrane Plasmique CH2 2022-2023Mbaye AbouPas encore d'évaluation
- CH3 - Cytosol-Cytosquelette - B.BDocument47 pagesCH3 - Cytosol-Cytosquelette - B.Bone asslanPas encore d'évaluation
- Technicien TerritorialDocument7 pagesTechnicien TerritorialPriscilliaArchenPas encore d'évaluation
- 1-Cours D IntroductionDocument40 pages1-Cours D Introductionml,zml,zPas encore d'évaluation
- Cours Membrane Plasmique & Hyaloplasme Microsoft Office WordDocument19 pagesCours Membrane Plasmique & Hyaloplasme Microsoft Office WordJoan Jean Jephthé Babo100% (4)
- Biochimie AlimentaireDocument63 pagesBiochimie AlimentaireCiobanu Vladislav100% (1)
- Cours 3. Membrane PlasmiqueDocument14 pagesCours 3. Membrane PlasmiqueNã BîlPas encore d'évaluation
- SVT - Fiche 1 - Organisation Fonctionnelle Du VivantDocument8 pagesSVT - Fiche 1 - Organisation Fonctionnelle Du VivantDjibril TourePas encore d'évaluation
- Adherence 2017Document26 pagesAdherence 2017laarabPas encore d'évaluation
- CytosqueletteDocument27 pagesCytosqueletteFatima BENHAFSSAPas encore d'évaluation
- Dissertation ScientifiqueDocument5 pagesDissertation Scientifiqueseydina123ababacarPas encore d'évaluation
- Synthèse La CelluleDocument6 pagesSynthèse La CelluleKamil LahbabiPas encore d'évaluation
- Chapitre 2 - Membrane Plasmique-Programme 2021 1Document9 pagesChapitre 2 - Membrane Plasmique-Programme 2021 1douniamhd096Pas encore d'évaluation
- RA19 Lycee G 1er ES T1 Membrane Plasmique V4 1172621Document9 pagesRA19 Lycee G 1er ES T1 Membrane Plasmique V4 1172621nouhailaelbouzidi879Pas encore d'évaluation
- Diapositives Fonctions Vtamines Et CoenzymesDocument12 pagesDiapositives Fonctions Vtamines Et CoenzymesAdmi BraccPas encore d'évaluation
- Synapses PDFDocument17 pagesSynapses PDFAdmi BraccPas encore d'évaluation
- Devenir Le Meilleur de Soi-Même - Abraham MaslowDocument497 pagesDevenir Le Meilleur de Soi-Même - Abraham MaslowKouakou Kouamé Rodrigue91% (11)
- Dup (01) Soc 180 (Questions - Reponses)Document8 pagesDup (01) Soc 180 (Questions - Reponses)Admi Bracc100% (1)
- La Cavitation Chez Les PlantesDocument9 pagesLa Cavitation Chez Les PlantesAdmi BraccPas encore d'évaluation
- SVT FASICULE 3eme 1Document56 pagesSVT FASICULE 3eme 1Methzo ThiamPas encore d'évaluation
- DIPLO - Dominance Et Propriétés PDFDocument4 pagesDIPLO - Dominance Et Propriétés PDFAdmi BraccPas encore d'évaluation
- Haplophase - Transmission Simultanée de Plus de Gènes PDFDocument2 pagesHaplophase - Transmission Simultanée de Plus de Gènes PDFAdmi BraccPas encore d'évaluation
- Pourquoi Active Windows Avce Kmspico Et Comment PDFDocument4 pagesPourquoi Active Windows Avce Kmspico Et Comment PDFAdmi BraccPas encore d'évaluation
- 2021 BOT262 CELL VEGET - CHAP3bis - Aout 09Document4 pages2021 BOT262 CELL VEGET - CHAP3bis - Aout 09Admi BraccPas encore d'évaluation
- Exercice 2 TD Haplophase PDFDocument1 pageExercice 2 TD Haplophase PDFAdmi BraccPas encore d'évaluation
- Chp2 Partie C.TDDocument2 pagesChp2 Partie C.TDAdmi BraccPas encore d'évaluation
- DIPLO - Intro Et Mono PDFDocument17 pagesDIPLO - Intro Et Mono PDFAdmi BraccPas encore d'évaluation
- Cas Des Ascomycètes PDFDocument3 pagesCas Des Ascomycètes PDFAdmi BraccPas encore d'évaluation
- Bot 134: Physiologie de La Nutrition Minérale Et de L'alimentation en Eau Chez Les VégétauxDocument4 pagesBot 134: Physiologie de La Nutrition Minérale Et de L'alimentation en Eau Chez Les VégétauxAdmi BraccPas encore d'évaluation
- Complementation PDFDocument5 pagesComplementation PDFAdmi BraccPas encore d'évaluation
- DIPLO Complementation PDFDocument4 pagesDIPLO Complementation PDFAdmi BraccPas encore d'évaluation
- Mise en Evidence, Symptomes Et Causes Des Carences en Elements MinerauxDocument9 pagesMise en Evidence, Symptomes Et Causes Des Carences en Elements MinerauxAdmi BraccPas encore d'évaluation
- Schema General de L'Absorption & Methodes D'EtudeDocument6 pagesSchema General de L'Absorption & Methodes D'EtudeAdmi BraccPas encore d'évaluation
- DIPLO - Epistasie Expliquée PDFDocument6 pagesDIPLO - Epistasie Expliquée PDFAdmi BraccPas encore d'évaluation
- Situation D'évaluation: ContexteDocument2 pagesSituation D'évaluation: ContexteAdmi BraccPas encore d'évaluation
- Epreuve de Physique Premiere C Annee 2020 Lycee Scientifique deDocument3 pagesEpreuve de Physique Premiere C Annee 2020 Lycee Scientifique deAdmi BraccPas encore d'évaluation
- Continu It ÉDocument150 pagesContinu It ÉPriscard100% (2)
- Caractéristiques Distinctives Entre Heredité Chromosomique Et Non Chromosomique PDFDocument2 pagesCaractéristiques Distinctives Entre Heredité Chromosomique Et Non Chromosomique PDFAdmi BraccPas encore d'évaluation
- Absorption de L'Eau Par La PlanteDocument8 pagesAbsorption de L'Eau Par La PlanteAdmi BraccPas encore d'évaluation
- Fiche Explicative de La Leçon - Caryotypes - NagwaDocument9 pagesFiche Explicative de La Leçon - Caryotypes - NagwaAdmi BraccPas encore d'évaluation
- Carto 6Document20 pagesCarto 6Admi BraccPas encore d'évaluation
- Bepc Blanc Régional 2023 Zone AboboDocument21 pagesBepc Blanc Régional 2023 Zone AboboAdmi Bracc100% (1)
- Haplophase - Différence Due À Un Seul Gène 1Document5 pagesHaplophase - Différence Due À Un Seul Gène 1Admi BraccPas encore d'évaluation
- Guide en Litterature Toute SerieDocument14 pagesGuide en Litterature Toute SerieAdmi BraccPas encore d'évaluation
- Les TaninsDocument2 pagesLes TaninsYuliya RomanteevaPas encore d'évaluation
- Les Trois CerveauxDocument5 pagesLes Trois CerveauxJean Claude EnriquePas encore d'évaluation
- BiocellulaireDocument4 pagesBiocellulairecarlio12Pas encore d'évaluation
- Mes RévisionsDocument22 pagesMes RévisionsPatience OnwinkpePas encore d'évaluation
- Cours Anatomie App Respiratoire, 2017Document66 pagesCours Anatomie App Respiratoire, 2017walid ouPas encore d'évaluation
- Medecine Humaine PDFDocument2 pagesMedecine Humaine PDFDiah BarbéPas encore d'évaluation
- Bac S - Sujet de SVT - Session 2019 - Emirats Arabes Unis: 1ère PARTIE: (8 Points)Document8 pagesBac S - Sujet de SVT - Session 2019 - Emirats Arabes Unis: 1ère PARTIE: (8 Points)Général Engongha MinangPas encore d'évaluation
- Anti InflammatoiresDocument6 pagesAnti InflammatoiresBENCHABANE OUSSAMAPas encore d'évaluation
- Epreuves de Résidanat Pharmacologie 2011 2016Document46 pagesEpreuves de Résidanat Pharmacologie 2011 2016Guzel MuzPas encore d'évaluation
- Cours RCPG-M1-2016-PCMDocument26 pagesCours RCPG-M1-2016-PCMIbrahimPas encore d'évaluation
- 2-Histologie de L'Appareil RespiratoireDocument61 pages2-Histologie de L'Appareil RespiratoireImene Imene100% (1)
- Récepteur de La Superfamille TNF AlphaDocument28 pagesRécepteur de La Superfamille TNF AlphaFatima ZahraPas encore d'évaluation
- Qu'advient-Il de Ces Milliards de Nanoparticules Dont Vous Eì Tes Devenu L'hoì Te ?Document11 pagesQu'advient-Il de Ces Milliards de Nanoparticules Dont Vous Eì Tes Devenu L'hoì Te ?GCo GroupPas encore d'évaluation
- La Posture de L Arbre PDFDocument61 pagesLa Posture de L Arbre PDFkoubyPas encore d'évaluation
- Les Antidiabteiques Oraux: Mécanisme D'actionDocument4 pagesLes Antidiabteiques Oraux: Mécanisme D'actioncao mengdePas encore d'évaluation
- Cours V La Réaction InflammatoireDocument14 pagesCours V La Réaction InflammatoiretinaPas encore d'évaluation
- Circulation Sanguine LDocument4 pagesCirculation Sanguine LDel FinePas encore d'évaluation
- 3 L'électrophysiologie CardiaqueDocument8 pages3 L'électrophysiologie Cardiaqueaya fifiPas encore d'évaluation
- Anémie MacrocytaireDocument7 pagesAnémie MacrocytaireMESSALI ABDELHAFID100% (1)
- Deshydratation 2Document3 pagesDeshydratation 2gagPas encore d'évaluation
- Synthèse Motifs D'hospitDocument3 pagesSynthèse Motifs D'hospitEnzo PiniPas encore d'évaluation
- Résumé SC Biologiques-1Document3 pagesRésumé SC Biologiques-1hamza malloukPas encore d'évaluation
- 4-Le Tissu ConjonctifDocument60 pages4-Le Tissu ConjonctifAy ManPas encore d'évaluation
- AnalyseDocument5 pagesAnalysegescomPas encore d'évaluation
- La Martingale Anatomie Physiologie Sémiologie Pour lEDN 2022Document358 pagesLa Martingale Anatomie Physiologie Sémiologie Pour lEDN 2022tinosan3Pas encore d'évaluation
- DR Mahdaoui Sources PDFDocument1 pageDR Mahdaoui Sources PDFKatia Hareb-Cherfaoui67% (3)
- Eudicot ArchaiquesDocument3 pagesEudicot ArchaiquesMar OuaPas encore d'évaluation
- TD Glucide Medecine 2020 2021 (Série)Document4 pagesTD Glucide Medecine 2020 2021 (Série)Sarah BelbachirPas encore d'évaluation
- Enzymologie Et BioénergétiqueDocument15 pagesEnzymologie Et BioénergétiqueZorba ZorbaPas encore d'évaluation
- Cours de Terminologie MédicaleDocument27 pagesCours de Terminologie MédicaleMohamedLashab0% (2)
- Manuel pour les débutants Fabriquez des savons naturelsD'EverandManuel pour les débutants Fabriquez des savons naturelsÉvaluation : 3 sur 5 étoiles3/5 (2)
- Encyclopédie de la magie des cristaux, des pierres précieuses et des métauxD'EverandEncyclopédie de la magie des cristaux, des pierres précieuses et des métauxÉvaluation : 4 sur 5 étoiles4/5 (116)
- Les matérialismes et la chimie: Perspectives philosophiques, historiques et scientifiquesD'EverandLes matérialismes et la chimie: Perspectives philosophiques, historiques et scientifiquesPas encore d'évaluation
- Lanthane et lanthanides: Les Grands Articles d'UniversalisD'EverandLanthane et lanthanides: Les Grands Articles d'UniversalisPas encore d'évaluation
- Macromolécules: Les Grands Articles d'UniversalisD'EverandMacromolécules: Les Grands Articles d'UniversalisPas encore d'évaluation
- Applications de la spectrophotomérie en phytochimie: sciencesD'EverandApplications de la spectrophotomérie en phytochimie: sciencesPas encore d'évaluation
- Chromatographie: Les Grands Articles d'UniversalisD'EverandChromatographie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Du Néant à la Formule Universelle et retour: La structure des particules élémentaires XIIIfD'EverandDu Néant à la Formule Universelle et retour: La structure des particules élémentaires XIIIfPas encore d'évaluation
- Equilibre Hormonal: Comment regagner l’équilibre d’hormones, la pulsion sexuelle, le sommeil et perdre du poids dès maintenantD'EverandEquilibre Hormonal: Comment regagner l’équilibre d’hormones, la pulsion sexuelle, le sommeil et perdre du poids dès maintenantPas encore d'évaluation
- Le Big Bang: Une Théorie Basée sur une Physique Invalide et des Mathématiques Erronées.D'EverandLe Big Bang: Une Théorie Basée sur une Physique Invalide et des Mathématiques Erronées.Pas encore d'évaluation
- Formation 3D par la pratique avec C#5 et WPF: Modeliser des moleculesD'EverandFormation 3D par la pratique avec C#5 et WPF: Modeliser des moleculesPas encore d'évaluation
- Stéréochimie organique: Les Grands Articles d'UniversalisD'EverandStéréochimie organique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Histoire de la chimie: Les Grands Articles d'UniversalisD'EverandHistoire de la chimie: Les Grands Articles d'UniversalisPas encore d'évaluation
- Vade-mecum du photographe: notice abrégée du daguerréotype et de la photographie sur papierD'EverandVade-mecum du photographe: notice abrégée du daguerréotype et de la photographie sur papierPas encore d'évaluation
- Équilibres en solution: Les Grands Articles d'UniversalisD'EverandÉquilibres en solution: Les Grands Articles d'UniversalisPas encore d'évaluation