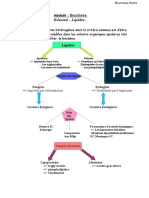
Vous aimerez peut-être aussi
- Glucides: Les Grands Articles d'UniversalisD'EverandGlucides: Les Grands Articles d'UniversalisPas encore d'évaluation
- Acides nucléiques: Les Grands Articles d'UniversalisD'EverandAcides nucléiques: Les Grands Articles d'UniversalisPas encore d'évaluation
- LipidesDocument42 pagesLipidesKEBIR ZAHRAPas encore d'évaluation
- Resume Les LipidesDocument14 pagesResume Les LipidesihcPas encore d'évaluation
- Lipides (Cours)Document14 pagesLipides (Cours)Rock In BandPas encore d'évaluation
- Chapitre 1-3 Les LipidesDocument17 pagesChapitre 1-3 Les Lipidesmohamadou moustapha gayePas encore d'évaluation
- 1 1 Structure Des Lipides PDFDocument12 pages1 1 Structure Des Lipides PDFNaouel BoudPas encore d'évaluation
- Cours Biochimie Lipides SV3Document65 pagesCours Biochimie Lipides SV3Zineb DahmaniPas encore d'évaluation
- Cours chimie bioorganique lipidesDocument6 pagesCours chimie bioorganique lipidesChaimaa asnounPas encore d'évaluation
- LIPIDES Biochimie StructuraleDocument82 pagesLIPIDES Biochimie StructuraleCheick SANOUPas encore d'évaluation
- BiochimieDocument59 pagesBiochimieAndréa Stennie RaoelimahefaPas encore d'évaluation
- 1.1. AG Structure, Propréités Et Méthodes de Séparation 2014Document22 pages1.1. AG Structure, Propréités Et Méthodes de Séparation 2014menzaouyedPas encore d'évaluation
- 2-Polycopie LipidesDocument27 pages2-Polycopie Lipidesyanis100% (1)
- Acides Gras Nomenclature Et Sources AlimentairesDocument8 pagesAcides Gras Nomenclature Et Sources AlimentairescoolsvpPas encore d'évaluation
- Les LipidesDocument25 pagesLes LipidesMamineTecDelmaPas encore d'évaluation
- Medecine Biochimie 1anDocument107 pagesMedecine Biochimie 1anhousam100% (2)
- Cours Biochimie Chapitre 2 Les LipidesDocument11 pagesCours Biochimie Chapitre 2 Les Lipidesazouzrouaida49Pas encore d'évaluation
- Cours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieDocument83 pagesCours - Biochimie Structurale - Lipides - CAG - UNA - 2019 - CopieStellaPas encore d'évaluation
- Structure Des Lipides Medecine DentaireDocument93 pagesStructure Des Lipides Medecine DentairezadiPas encore d'évaluation
- LipideDocument22 pagesLipideKarima AarabePas encore d'évaluation
- UE1 - Biochimie - Les GlucidesDocument71 pagesUE1 - Biochimie - Les Glucidessun-nee-chan9Pas encore d'évaluation
- Présentation 3Document32 pagesPrésentation 3Yahia BerbouchiPas encore d'évaluation
- COURS LIPIDES FMS 2022 - 2023 (Pr. SAID)Document19 pagesCOURS LIPIDES FMS 2022 - 2023 (Pr. SAID)Abdelhamid ArouiPas encore d'évaluation
- Biochimie - Riad BenchouchaDocument107 pagesBiochimie - Riad Benchouchakarim islamPas encore d'évaluation
- Les LipidesDocument8 pagesLes Lipidesyousraghanem23Pas encore d'évaluation
- ACIDE GRAS_ etudiantsDocument48 pagesACIDE GRAS_ etudiantsZaki ByzPas encore d'évaluation
- Biochimie Lipide PDF, ManuscritDocument8 pagesBiochimie Lipide PDF, Manuscritlaoudi sarahPas encore d'évaluation
- LIPIDES Biochimie Structurale-1Document69 pagesLIPIDES Biochimie Structurale-1Cheick SANOUPas encore d'évaluation
- Cours de Biochimie Structurale 2022 2023 LIPIDEDocument25 pagesCours de Biochimie Structurale 2022 2023 LIPIDEvivaspro900Pas encore d'évaluation
- Seance 5Document17 pagesSeance 5Pascal GadedjissoPas encore d'évaluation
- Lip IdesDocument8 pagesLip IdesYousra INDIAPas encore d'évaluation
- 1-Caracteres Generaux Des LipidesDocument65 pages1-Caracteres Generaux Des Lipidesbubble townPas encore d'évaluation
- Biochimie 01 - Biomolécules Constitutives Des Cellules Glucides-LipidesDocument22 pagesBiochimie 01 - Biomolécules Constitutives Des Cellules Glucides-LipidesNathan CohenPas encore d'évaluation
- Biochimie Structurale ESTBADocument79 pagesBiochimie Structurale ESTBAtolisso.yawoviPas encore d'évaluation
- 1-Structure Des OsesDocument25 pages1-Structure Des Oseslara milanoPas encore d'évaluation
- UE1 Biochimie Glucides 3 CoursDocument67 pagesUE1 Biochimie Glucides 3 Courssun-nee-chan9Pas encore d'évaluation
- 2-Résumé Bournisa Amira 2015-2016Document25 pages2-Résumé Bournisa Amira 2015-2016Farouk DjenienPas encore d'évaluation
- 1 - Les LipidesDocument25 pages1 - Les LipidesFanny GrouxPas encore d'évaluation
- Les LipidesDocument32 pagesLes LipidesWã RdäPas encore d'évaluation
- Lip IdesDocument7 pagesLip IdesMouna Boujelben NejiPas encore d'évaluation
- Cours Sur Les LipidesDocument13 pagesCours Sur Les LipidesbonobogeantPas encore d'évaluation
- Cours Glucides.2021-22.doc Copies EtudiantsDocument31 pagesCours Glucides.2021-22.doc Copies Etudiantsimad essaidyPas encore d'évaluation
- Cours Biochimie ST Lipides Alouani-Séance 6 PDFDocument17 pagesCours Biochimie ST Lipides Alouani-Séance 6 PDFBushraPas encore d'évaluation
- Glucides 3lbiol 2021Document71 pagesGlucides 3lbiol 2021KEBIR ZAHRAPas encore d'évaluation
- Chap 1 Les Glucides Structure Et MétabolismeDocument75 pagesChap 1 Les Glucides Structure Et Métabolismeafaf gou100% (1)
- Cours Glucides 2021 2022Document27 pagesCours Glucides 2021 2022Abdelhamid ArouiPas encore d'évaluation
- Cours Lipides 20 21Document16 pagesCours Lipides 20 21oumayma salihiPas encore d'évaluation
- LES LIPIDES Sructure Et Métabolque 2010Document162 pagesLES LIPIDES Sructure Et Métabolque 2010Lyes DahmaniPas encore d'évaluation
- Les GlucidesDocument40 pagesLes GlucidesRadhia KossaiPas encore d'évaluation
- Matière Vivante-1Document53 pagesMatière Vivante-1malomedPas encore d'évaluation
- Biochimie Structurale Des Acides Gras UIL-1Document53 pagesBiochimie Structurale Des Acides Gras UIL-1Ay ManPas encore d'évaluation
- 01 - structures des lipides et AGGDocument9 pages01 - structures des lipides et AGGeladib p1Pas encore d'évaluation
- Lipides Métabolisme Poly LaresbaDocument119 pagesLipides Métabolisme Poly LaresbaFrançois regis NikièmaPas encore d'évaluation
- Les Corps Gras AlimentairesDocument18 pagesLes Corps Gras AlimentairesZineb BelalaPas encore d'évaluation
- Lipides Cours 01 PDFDocument26 pagesLipides Cours 01 PDFpublicité en flopOfficielPas encore d'évaluation
- LipideDocument28 pagesLipideChanez FekirPas encore d'évaluation
- Biochimie FDocument7 pagesBiochimie FMathin BrouPas encore d'évaluation
- Chap.2 Les LipidesDocument2 pagesChap.2 Les Lipidesangiehayek1234Pas encore d'évaluation
- Protéines & Acides Aminés (Cours)Document10 pagesProtéines & Acides Aminés (Cours)Rock In BandPas encore d'évaluation
- Vitamines (Cours)Document11 pagesVitamines (Cours)Rock In BandPas encore d'évaluation
- Correction D'annales Physio/BiophysiqueDocument19 pagesCorrection D'annales Physio/Biophysiquesun-nee-chan9Pas encore d'évaluation
- Concours Blanc Droit/Eco/Socio/Psycho SujetDocument19 pagesConcours Blanc Droit/Eco/Socio/Psycho Sujetsun-nee-chan9Pas encore d'évaluation
- Cours Maths de Première - Généralités Sur Les FonctionsDocument3 pagesCours Maths de Première - Généralités Sur Les Fonctionssun-nee-chan9Pas encore d'évaluation
- Concours Blanc Droit/Eco/Socio/Psycho CorrectionDocument20 pagesConcours Blanc Droit/Eco/Socio/Psycho Correctionsun-nee-chan9Pas encore d'évaluation
- EXO REVISION HISTO p1Document11 pagesEXO REVISION HISTO p1sun-nee-chan9Pas encore d'évaluation
- CB Sujet Biocell/Embryo/HistoDocument21 pagesCB Sujet Biocell/Embryo/Histosun-nee-chan9Pas encore d'évaluation
- UE2 Embryologie - Gamétogenèse (Final)Document18 pagesUE2 Embryologie - Gamétogenèse (Final)sun-nee-chan9Pas encore d'évaluation
- UE7-socio-5-inégalités Sociales de Santé (ACC)Document12 pagesUE7-socio-5-inégalités Sociales de Santé (ACC)sun-nee-chan9Pas encore d'évaluation
- UE2 - Histologie - Tissu NerveuxDocument38 pagesUE2 - Histologie - Tissu Nerveuxsun-nee-chan9Pas encore d'évaluation
- UE3-physique-1-notions de BaseDocument26 pagesUE3-physique-1-notions de Basesun-nee-chan9Pas encore d'évaluation
- UE2-Embryo-3-Fécondation (Final)Document22 pagesUE2-Embryo-3-Fécondation (Final)sun-nee-chan9Pas encore d'évaluation
- UE3 - Physique - Radiobiologie Et RadioprotectionDocument7 pagesUE3 - Physique - Radiobiologie Et Radioprotectionsun-nee-chan9Pas encore d'évaluation
- Cours Maths de Première - Quelques Définitions de Notions de Bases en MathsDocument3 pagesCours Maths de Première - Quelques Définitions de Notions de Bases en Mathssun-nee-chan9Pas encore d'évaluation
- UE7 - Economie - 2 - Croissance Et DéveloppementDocument10 pagesUE7 - Economie - 2 - Croissance Et Développementsun-nee-chan9Pas encore d'évaluation
- UE2-Embryo-1-Méïose FinalDocument6 pagesUE2-Embryo-1-Méïose Finalsun-nee-chan9Pas encore d'évaluation
- UE7-Sociologie - 6 - Emploi Et TravailDocument23 pagesUE7-Sociologie - 6 - Emploi Et Travailsun-nee-chan9Pas encore d'évaluation
- UE2 Embryologie - Gamétogenèse (Final)Document18 pagesUE2 Embryologie - Gamétogenèse (Final)sun-nee-chan9Pas encore d'évaluation
- UE2-intégration - 14-Différenciation CellulaireDocument11 pagesUE2-intégration - 14-Différenciation Cellulairesun-nee-chan9Pas encore d'évaluation
- UE2 La-Membrane-PlasmiqueDocument17 pagesUE2 La-Membrane-Plasmiquesun-nee-chan9Pas encore d'évaluation
- UE2 Embryologie - Les 2e Et 3e Semaines de Développement (Final)Document16 pagesUE2 Embryologie - Les 2e Et 3e Semaines de Développement (Final)sun-nee-chan9Pas encore d'évaluation
- UE2 - Biologie Cellulaire - Production D - Énergie Dans La CelluleDocument16 pagesUE2 - Biologie Cellulaire - Production D - Énergie Dans La Cellulesun-nee-chan9Pas encore d'évaluation
- UE2 - Biologie Cellulaire - Le CytosolDocument11 pagesUE2 - Biologie Cellulaire - Le Cytosolsun-nee-chan9Pas encore d'évaluation
- UE1-Chimie-9-fonctions DivalentesDocument21 pagesUE1-Chimie-9-fonctions Divalentessun-nee-chan9Pas encore d'évaluation
- UE1 Chimie 1 AtomistimeDocument29 pagesUE1 Chimie 1 Atomistimesun-nee-chan9Pas encore d'évaluation
- UE1-Chimie-4-effets ÉlectroniquesDocument14 pagesUE1-Chimie-4-effets Électroniquessun-nee-chan9Pas encore d'évaluation
- UE1 Biochimie Glucides 3 CoursDocument67 pagesUE1 Biochimie Glucides 3 Courssun-nee-chan9Pas encore d'évaluation
- UE2 - Biologie Cellulaire - Molécule de Surface Et D - AdhérenceDocument19 pagesUE2 - Biologie Cellulaire - Molécule de Surface Et D - Adhérencesun-nee-chan9Pas encore d'évaluation
- UE1 Biochimie 6 - Structure Du GénomeDocument18 pagesUE1 Biochimie 6 - Structure Du Génomesun-nee-chan9Pas encore d'évaluation
- EMD 2 Biochimie 2023Document4 pagesEMD 2 Biochimie 2023ikou ikramPas encore d'évaluation
- Introduction Sur Le Métabolisme LipidesDocument21 pagesIntroduction Sur Le Métabolisme Lipidescep100% (1)
- Ig FinalDocument13 pagesIg FinalDr VerdasPas encore d'évaluation
- Constituantsmatiere VivanteDocument3 pagesConstituantsmatiere VivanteFatma Sy100% (1)
- Technologie de Poissons Et DérivésDocument64 pagesTechnologie de Poissons Et DérivésPeti' Pou89% (9)
- Dyslipidemies Nouvelles Therapeutiques 2006Document30 pagesDyslipidemies Nouvelles Therapeutiques 2006megenniPas encore d'évaluation
- Les Principales Propriétés Physico Chimiques Des Constituants Alimentaires-Converti 2Document5 pagesLes Principales Propriétés Physico Chimiques Des Constituants Alimentaires-Converti 2Marwa BachirPas encore d'évaluation
- ED3 GluLip 20007-08Document13 pagesED3 GluLip 20007-08AissaPas encore d'évaluation
- Les Maladies Metaboliques Chez Les Ovins: Ecole Nationale Veterinaire D'AlfortDocument140 pagesLes Maladies Metaboliques Chez Les Ovins: Ecole Nationale Veterinaire D'Alfortهشام شريفيPas encore d'évaluation
- Cours Lipides 2 AG1 V 2023Document37 pagesCours Lipides 2 AG1 V 2023Amani Mann'fangan EphremePas encore d'évaluation
- Chirurgie Digestive2Document151 pagesChirurgie Digestive2Père ProsperPas encore d'évaluation
- CHAP13 - Relations Avec L'environnementDocument11 pagesCHAP13 - Relations Avec L'environnementmadaulePas encore d'évaluation
- Serie N°3 - Correction Structure Et Métabolisme Des LipidesDocument3 pagesSerie N°3 - Correction Structure Et Métabolisme Des Lipidesafaf gou100% (2)
- Nutrition VégétaleDocument16 pagesNutrition VégétaleVegan FuturePas encore d'évaluation
- Fiche de TP de BiochimieDocument29 pagesFiche de TP de BiochimieChristian NgalaPas encore d'évaluation
- Nutrition Chez Les Athletes-2Document37 pagesNutrition Chez Les Athletes-2christellePas encore d'évaluation
- (CHIEN) Tableau Simplifié - Besoins Du ChiensDocument1 page(CHIEN) Tableau Simplifié - Besoins Du Chiensar_scribdcomPas encore d'évaluation
- Bi102 LipidesDocument112 pagesBi102 LipidesMLAN HesnaPas encore d'évaluation
- Intro Et MPDocument22 pagesIntro Et MPWissam TizaPas encore d'évaluation
- Recueil Classement Tarifaire 2010Document66 pagesRecueil Classement Tarifaire 2010belelou2010Pas encore d'évaluation
- (Glucide-Lipide PR KABBAJDocument165 pages(Glucide-Lipide PR KABBAJskeleton93Pas encore d'évaluation
- These UTC Aurelien DagmeyDocument8 pagesThese UTC Aurelien Dagmeynaomiesolefack03Pas encore d'évaluation
- Structure BactérienneDocument4 pagesStructure BactérienneGHERMI .MPas encore d'évaluation
- Questions de Lexamen 1Document39 pagesQuestions de Lexamen 1dhouha14ghorbelPas encore d'évaluation
- 21 Calcule de La Valeur ÉnergétiqueDocument3 pages21 Calcule de La Valeur ÉnergétiquedzsergioPas encore d'évaluation
- Notes de Cours Biol 1133 1143Document156 pagesNotes de Cours Biol 1133 1143DL3 ProjectPas encore d'évaluation
- Bases de La NutritionDocument12 pagesBases de La Nutritionhicham lekfifiPas encore d'évaluation
- Notions Fondamentales Sur Corps GrasDocument14 pagesNotions Fondamentales Sur Corps GrasRayène BekirPas encore d'évaluation
- Composition Chimique Des Aliments Du BetailDocument40 pagesComposition Chimique Des Aliments Du BetailMohamed Habib Brinsi100% (1)
- Syllqbus Bilogie Cliniaue PDFDocument352 pagesSyllqbus Bilogie Cliniaue PDFEmmanuel100% (2)